

Dynamic landscapes of four histone modifications during deetiolation in Arabidopsis
- PMID: 20008096
- PMCID: PMC2814509
- DOI: 10.1105/tpc.109.066845
Dynamic landscapes of four histone modifications during deetiolation in Arabidopsis
Abstract
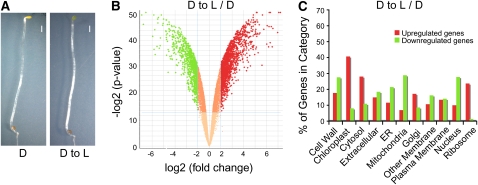
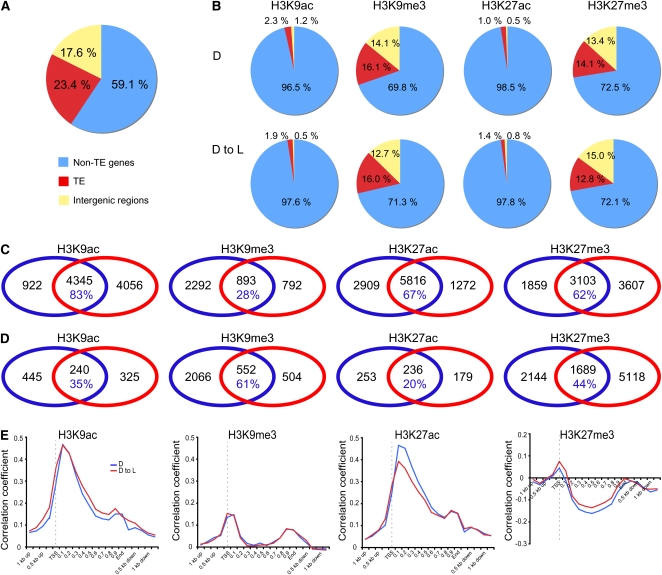
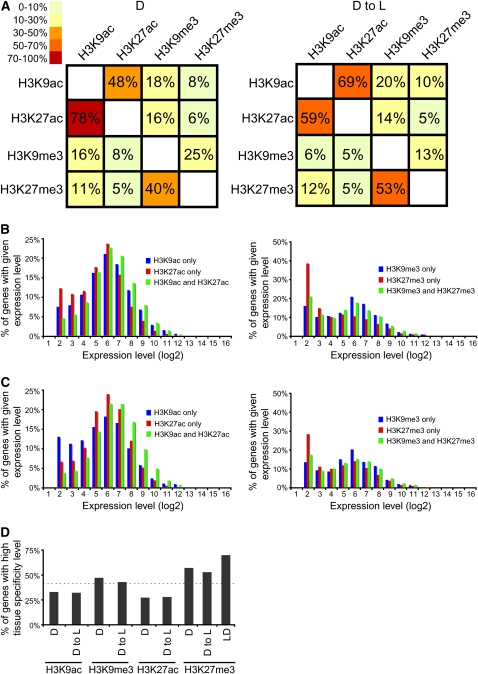
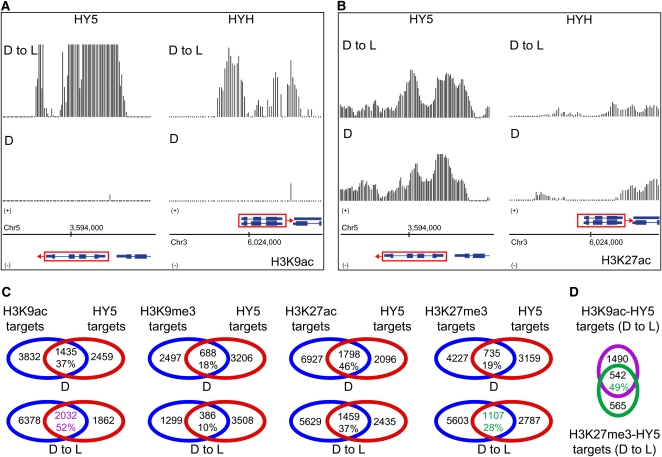
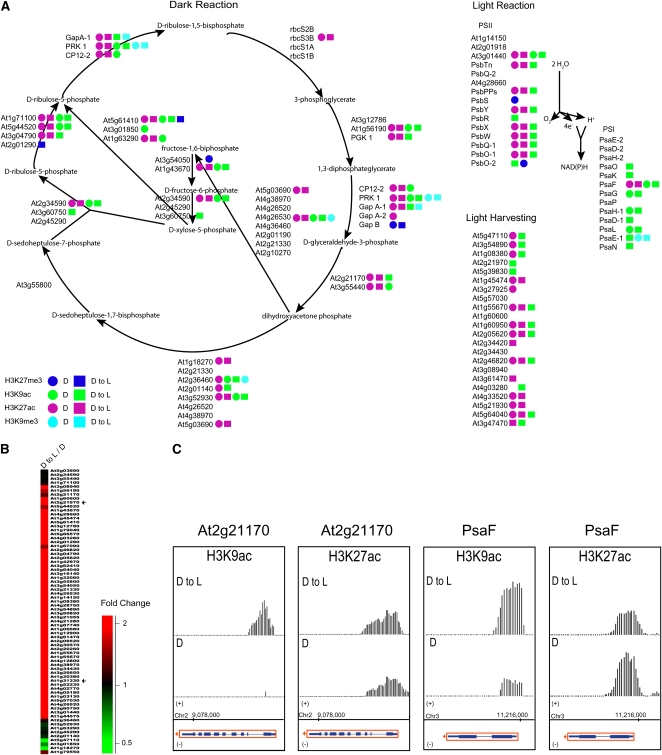
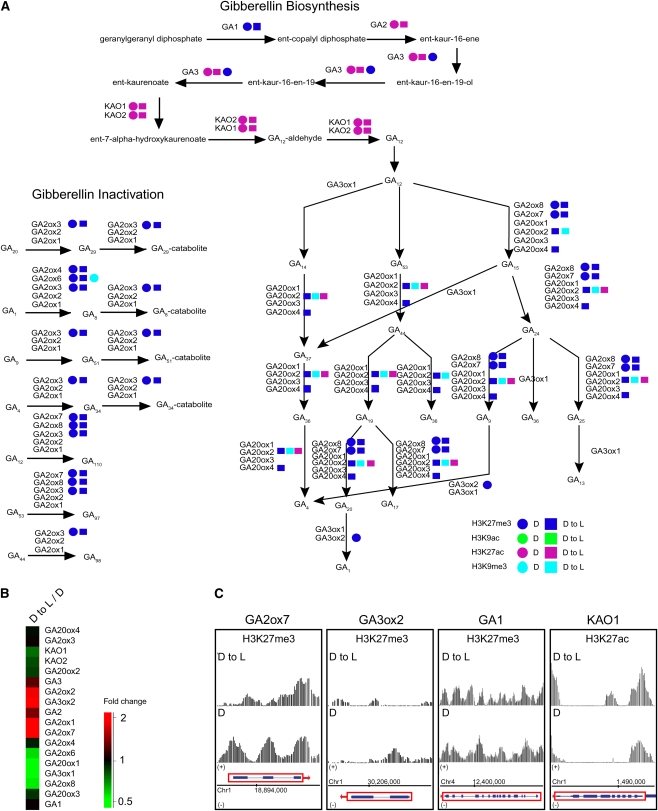
Although landscapes of several histone marks are now available for Arabidopsis thaliana and Oryza sativa, such profiles remain static and do not provide information about dynamic changes of plant epigenomes in response to developmental or environmental cues. Here, we analyzed the effects of light on four histone modifications (acetylation and trimethylation of lysines 9 and 27 on histone H3: H3K9ac, H3K9me3, H3K27ac, and H3K27me3, respectively). Our genome-wide profiling of H3K9ac and H3K27ac revealed that these modifications are nontransposable element gene-specific. By contrast, we found that H3K9me3 and H3K27me3 target nontransposable element genes, but also intergenic regions and transposable elements. Specific light conditions affected the number of modified regions as well as the overall correlation strength between the presence of specific modifications and transcription. Furthermore, we observed that acetylation marks not only ELONGATED HYPOCOTYL5 and HY5-HOMOLOG upon deetiolation, but also their downstream targets. We found that the activation of photosynthetic genes correlates with dynamic acetylation changes in response to light, while H3K27ac and H3K27me3 potentially contribute to light regulation of the gibberellin metabolism. Thus, this work provides a dynamic portrait of the variations in histone modifications in response to the plant's changing light environment and strengthens the concept that histone modifications represent an additional layer of control for light-regulated genes involved in photomorphogenesis.
Figures

Similar articles
-
Genome-wide profiling of histone H3 lysine 9 acetylation and dimethylation in Arabidopsis reveals correlation between multiple histone marks and gene expression.Plant Mol Biol. 2010 Apr;72(6):585-95. doi: 10.1007/s11103-009-9594-7. Plant Mol Biol. 2010. PMID: 20054610
-
Histone modifications and expression of light-regulated genes in Arabidopsis are cooperatively influenced by changing light conditions.Plant Physiol. 2008 Aug;147(4):2070-83. doi: 10.1104/pp.108.122929. Epub 2008 Jun 11. Plant Physiol. 2008. PMID: 18550682 Free PMC article.
-
Ethylene induces combinatorial effects of histone H3 acetylation in gene expression in Arabidopsis.BMC Genomics. 2017 Jul 17;18(1):538. doi: 10.1186/s12864-017-3929-6. BMC Genomics. 2017. PMID: 28716006 Free PMC article.
-
Epigenomics in stress tolerance of plants under the climate change.Mol Biol Rep. 2023 Jul;50(7):6201-6216. doi: 10.1007/s11033-023-08539-6. Epub 2023 Jun 9. Mol Biol Rep. 2023. PMID: 37294468 Review.
-
Histone modifications in transcriptional activation during plant development.Biochim Biophys Acta. 2011 Oct;1809(10):567-76. doi: 10.1016/j.bbagrm.2011.07.001. Epub 2011 Jul 14. Biochim Biophys Acta. 2011. PMID: 21777708 Review.
Cited by
-
Regulation of transcription by the Arabidopsis UVR8 photoreceptor involves a specific histone modification.Plant Mol Biol. 2016 Nov;92(4-5):425-443. doi: 10.1007/s11103-016-0522-3. Epub 2016 Aug 17. Plant Mol Biol. 2016. PMID: 27534420 Free PMC article.
-
H4 acetylation by the NuA4 complex is required for plastid transcription and chloroplast biogenesis.Nat Plants. 2022 Sep;8(9):1052-1063. doi: 10.1038/s41477-022-01229-4. Epub 2022 Aug 29. Nat Plants. 2022. PMID: 36038656
-
Tissue-specific transcriptomic analysis uncovers potential roles of natural antisense transcripts in Arabidopsis heat stress response.Front Plant Sci. 2022 Sep 8;13:997967. doi: 10.3389/fpls.2022.997967. eCollection 2022. Front Plant Sci. 2022. PMID: 36160979 Free PMC article.
-
Structural basis of a histone H3 lysine 4 demethylase required for stem elongation in rice.PLoS Genet. 2013;9(1):e1003239. doi: 10.1371/journal.pgen.1003239. Epub 2013 Jan 24. PLoS Genet. 2013. PMID: 23357881 Free PMC article.
-
Genome-wide identification of long noncoding natural antisense transcripts and their responses to light in Arabidopsis.Genome Res. 2014 Mar;24(3):444-53. doi: 10.1101/gr.165555.113. Epub 2014 Jan 8. Genome Res. 2014. PMID: 24402519 Free PMC article.
References
-
- Alabadi, D., Gallego-Bartolome, J., Orlando, L., Garcia-Carcel, L., Rubio, V., Martinez, C., Frigerio, M., Iglesias-Pedraz, J.M., Espinosa, A., Deng, X.W., and Blazquez, M.A. (2008). Gibberellins modulate light signaling pathways to prevent Arabidopsis seedling de-etiolation in darkness. Plant J. 53 324–335. - PubMed
-
- Benhamed, M., et al. (2008). Genome-scale Arabidopsis promoter array identifies targets of the histone acetyltransferase GCN5. Plant J. 56 493–504. - PubMed
-
- Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc., B 57 289–300.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
