

Quantitation of cellular deoxynucleoside triphosphates
- PMID: 20008099
- PMCID: PMC2847218
- DOI: 10.1093/nar/gkp1141
Quantitation of cellular deoxynucleoside triphosphates
Abstract
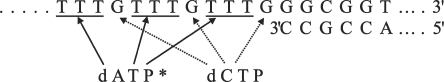
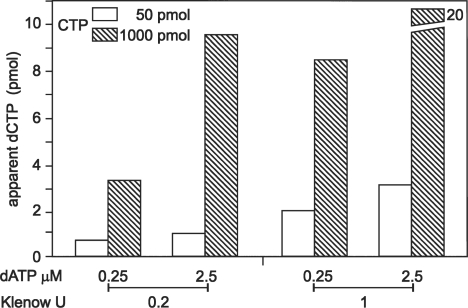
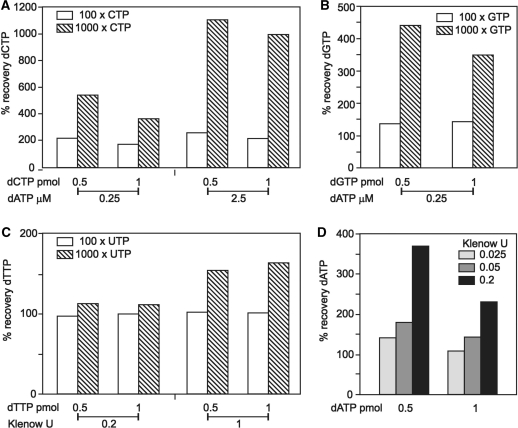
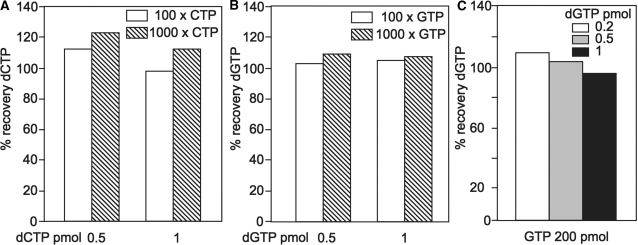
Eukaryotic cells contain a delicate balance of minute amounts of the four deoxyribonucleoside triphosphates (dNTPs), sufficient only for a few minutes of DNA replication. Both a deficiency and a surplus of a single dNTP may result in increased mutation rates, faulty DNA repair or mitochondrial DNA depletion. dNTPs are usually quantified by an enzymatic assay in which incorporation of radioactive dATP (or radioactive dTTP in the assay for dATP) into specific synthetic oligonucleotides by a DNA polymerase is proportional to the concentration of the unknown dNTP. We find that the commonly used Klenow DNA polymerase may substitute the corresponding ribonucleotide for the unknown dNTP leading in some instances to a large overestimation of dNTPs. We now describe assay conditions for each dNTP that avoid ribonucleotide incorporation. For the dTTP and dATP assays it suffices to minimize the concentrations of the Klenow enzyme and of labeled dATP (or dTTP); for dCTP and dGTP we had to replace the Klenow enzyme with either the Taq DNA polymerase or Thermo Sequenase. We suggest that in some earlier reports ribonucleotide incorporation may have caused too high values for dGTP and dCTP.
Figures
References
-
- Meuth M. The molecular basis of mutations induced by deoxyribonucleoside triphosphate pool imbalances in mammalian cells. Exp.Cell Res. 1989;181:305–316. - PubMed
-
- Reichard P. Interactions between deoxyribonucleotide and DNA synthesis (1988) Annu. Rev. Biochem. 1988;57:349–374. - PubMed
-
- Kunz BA, Kohalmi SE, Kunkel TA, Mathews CK, MacIntosh EM, Reidy IA. Deoxyribonucleoside triphosphate levels: a critical factor in the maintenance of genetic stability. Mutat. Res. 1994;318:1–64. - PubMed
-
- Spinazzola A, Zeviani M. Disorders of nuclear-mitochondrial intergenomic signaling. Gene. 2005;354:162–168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
