

Targeting distinct tumor-infiltrating myeloid cells by inhibiting CSF-1 receptor: combating tumor evasion of antiangiogenic therapy
- PMID: 20008303
- PMCID: PMC2826767
- DOI: 10.1182/blood-2009-08-237412
Targeting distinct tumor-infiltrating myeloid cells by inhibiting CSF-1 receptor: combating tumor evasion of antiangiogenic therapy
Abstract
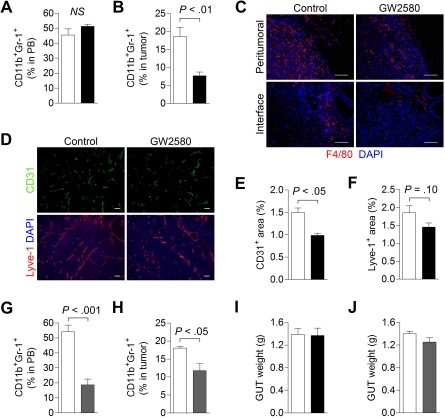
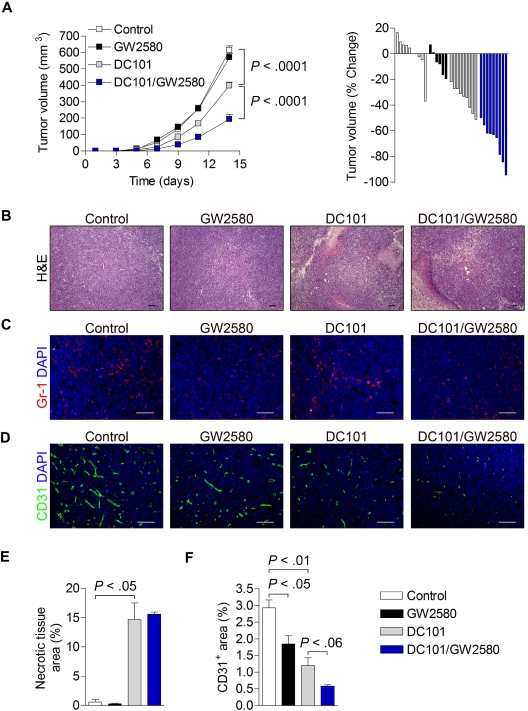
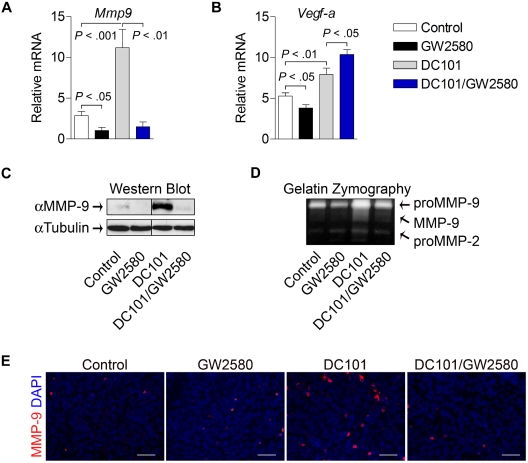
Tumor-infiltrating myeloid cells (TIMs) support tumor growth by promoting angiogenesis and suppressing antitumor immune responses. CSF-1 receptor (CSF1R) signaling is important for the recruitment of CD11b(+)F4/80(+) tumor-associated macrophages (TAMs) and contributes to myeloid cell-mediated angiogenesis. However, the impact of the CSF1R signaling pathway on other TIM subsets, including CD11b(+)Gr-1(+) myeloid-derived suppressor cells (MDSCs), is unknown. Tumor-infiltrating MDSCs have also been shown to contribute to tumor angiogenesis and have recently been implicated in tumor resistance to antiangiogenic therapy, yet their precise involvement in these processes is not well understood. Here, we use the selective pharmacologic inhibitor of CSF1R signaling, GW2580, to demonstrate that CSF-1 regulates the tumor recruitment of CD11b(+)Gr-1(lo)Ly6C(hi) mononuclear MDSCs. Targeting these TIM subsets inhibits tumor angiogenesis associated with reduced expression of proangiogenic and immunosuppressive genes. Combination therapy using GW2580 with an anti-VEGFR-2 antibody synergistically suppresses tumor growth and severely impairs tumor angiogenesis along with reverting at least one TIM-mediated antiangiogenic compensatory mechanism involving MMP-9. These data highlight the importance of CSF1R signaling in the recruitment and function of distinct TIM subsets, including MDSCs, and validate the benefits of targeting CSF1R signaling in combination with antiangiogenic drugs for the treatment of solid cancers.
Figures
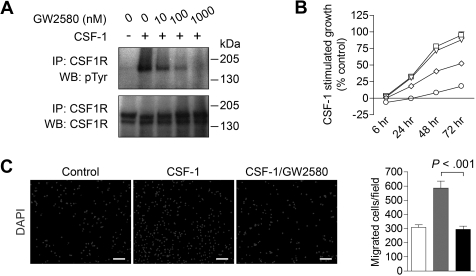
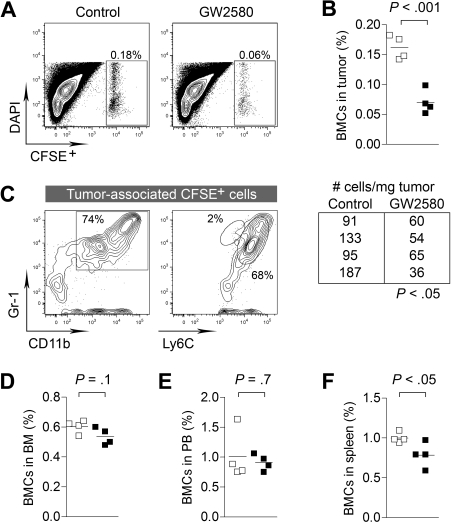
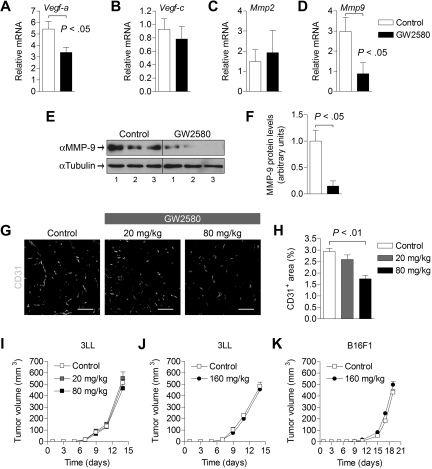
 , CSF-1; ■, CSF-1/GW2580). Scale bar represents 100 μm.
, CSF-1; ■, CSF-1/GW2580). Scale bar represents 100 μm.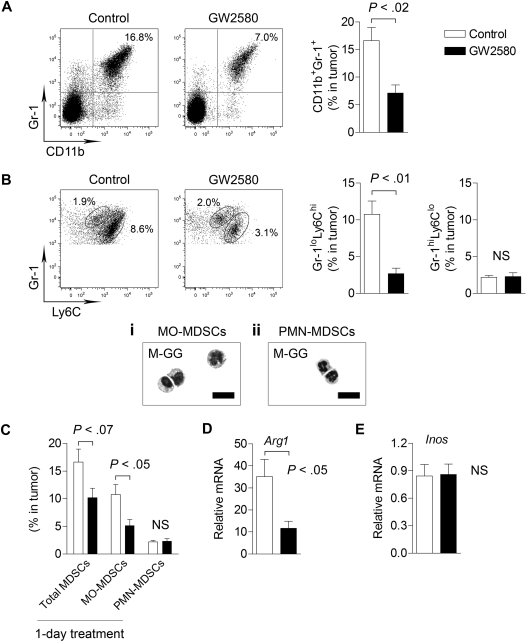
References
-
- Murdoch C, Muthana M, Coffelt SB, et al. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer. 2008;8(8):618–631. - PubMed
-
- Lewis CE, Pollard JW. Distinct role of macrophages in different tumor microenvironments. Cancer Res. 2006;66(2):605–612. - PubMed
-
- Gordon S. Alternative activation of macrophages. Nat Rev Immunol. 2003;3(1):23–35. - PubMed
-
- Sawanobori Y, Ueha S, Kurachi M, et al. Chemokine-mediated rapid turnover of myeloid-derived suppressor cells in tumor-bearing mice. Blood. 2008;111(12):5457–5466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
