

Regulatory role of the MisR/S two-component system in hemoglobin utilization in Neisseria meningitidis
- PMID: 20008531
- PMCID: PMC2825934
- DOI: 10.1128/IAI.00363-09
Regulatory role of the MisR/S two-component system in hemoglobin utilization in Neisseria meningitidis
Abstract
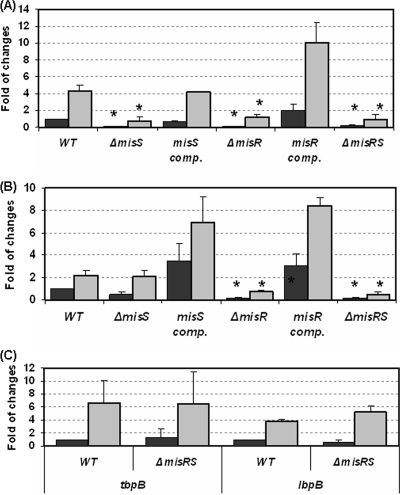
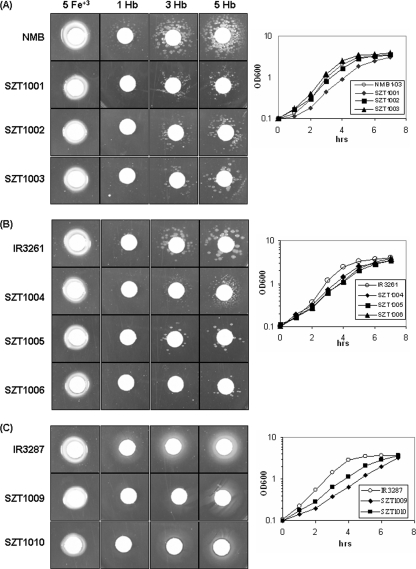
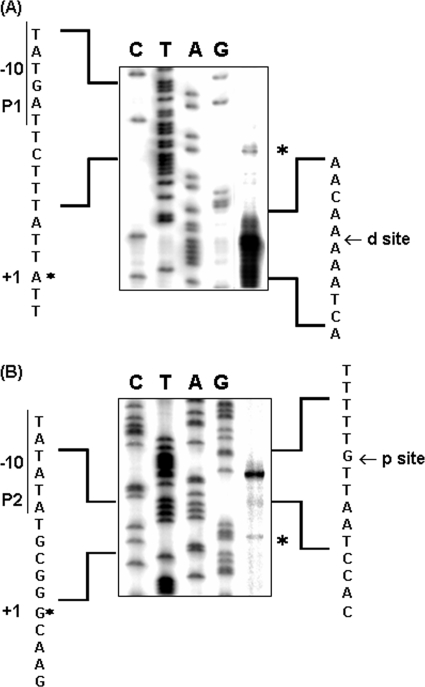
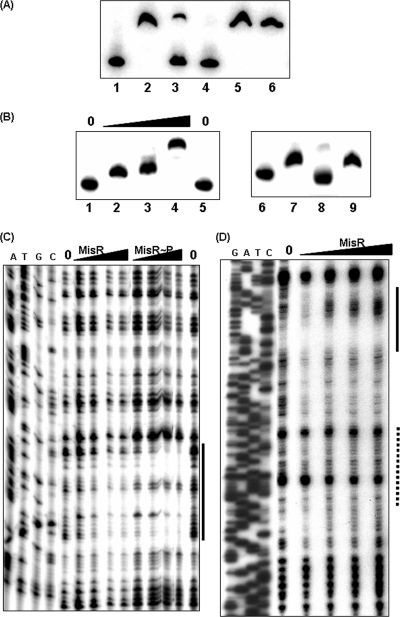
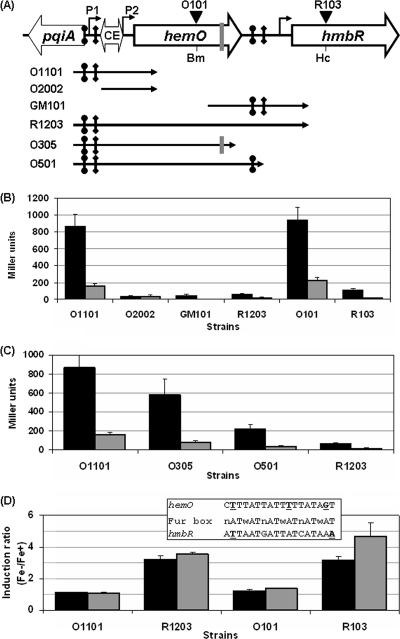
Outer membrane iron receptors are some of the major surface entities that are critical for meningococcal pathogenesis. The gene encoding the meningococcal hemoglobin receptor, HmbR, is both independently transcribed and transcriptionally linked to the upstream gene hemO, which encodes a heme oxygenase. The MisR/S two-component system was previously determined to regulate hmbR transcription, and its hemO and hmbR regulatory mechanisms were characterized further here. The expression of hemO and hmbR was downregulated in misR/S mutants under both iron-replete and iron-restricted conditions, and the downregulation could be reversed by complementation. No significant changes in expression of other iron receptors were detected, suggesting that the MisR/S system specifically regulates hmbR. When hemoglobin was the sole iron source, growth defects were detected in the mutants. Primer extension analysis identified a promoter upstream of the hemO-associated Correia element (CE) and another promoter at the proximal end of CE, and processed transcripts previously identified for other cotranscribed CEs were also detected, suggesting that there may be posttranscriptional regulation. MisR directly interacts with sequences upstream of the CE and upstream of the hmbR Fur binding site and thus independently regulates hemO and hmbR. Analysis of transcriptional reporters of hemO and hmbR further demonstrated the positive role of the MisR/S system and showed that the transcription of hmbR initiated from hemO was significantly reduced. A comparison of the effects of the misS mutation under iron-replete and iron-depleted conditions suggested that activation by the MisR/S system and iron-mediated repression by Fur act independently. Thus, the expression of hemO and hmbR is coordinately controlled by multiple independent regulatory mechanisms, including the MisR/S two-component system.
Figures

Similar articles
-
Use of heme compounds as iron sources by pathogenic neisseriae requires the product of the hemO gene.J Bacteriol. 2000 Jan;182(2):439-47. doi: 10.1128/JB.182.2.439-447.2000. J Bacteriol. 2000. PMID: 10629191 Free PMC article.
-
HmbR outer membrane receptors of pathogenic Neisseria spp.: iron-regulated, hemoglobin-binding proteins with a high level of primary structure conservation.J Bacteriol. 1996 Aug;178(15):4670-8. doi: 10.1128/jb.178.15.4670-4678.1996. J Bacteriol. 1996. PMID: 8755899 Free PMC article.
-
MisR/MisS two-component regulon in Neisseria meningitidis.Infect Immun. 2008 Feb;76(2):704-16. doi: 10.1128/IAI.01007-07. Epub 2007 Dec 3. Infect Immun. 2008. PMID: 18056476 Free PMC article.
-
Iron transport systems in Neisseria meningitidis.Microbiol Mol Biol Rev. 2004 Mar;68(1):154-71. doi: 10.1128/MMBR.68.1.154-171.2004. Microbiol Mol Biol Rev. 2004. PMID: 15007100 Free PMC article. Review.
-
Fur-mediated global regulatory circuits in pathogenic Neisseria species.J Bacteriol. 2012 Dec;194(23):6372-81. doi: 10.1128/JB.00262-12. Epub 2012 Aug 10. J Bacteriol. 2012. PMID: 22885296 Free PMC article. Review.
Cited by
-
The genes that encode the gonococcal transferrin binding proteins, TbpB and TbpA, are differentially regulated by MisR under iron-replete and iron-depleted conditions.Mol Microbiol. 2016 Oct;102(1):137-51. doi: 10.1111/mmi.13450. Epub 2016 Jul 18. Mol Microbiol. 2016. PMID: 27353397 Free PMC article.
-
The MisR Response Regulator Is Necessary for Intrinsic Cationic Antimicrobial Peptide and Aminoglycoside Resistance in Neisseria gonorrhoeae.Antimicrob Agents Chemother. 2016 Jul 22;60(8):4690-700. doi: 10.1128/AAC.00823-16. Print 2016 Aug. Antimicrob Agents Chemother. 2016. PMID: 27216061 Free PMC article.
-
A comparison of the endotoxin biosynthesis and protein oxidation pathways in the biogenesis of the outer membrane of Escherichia coli and Neisseria meningitidis.Front Cell Infect Microbiol. 2012 Dec 20;2:162. doi: 10.3389/fcimb.2012.00162. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 23267440 Free PMC article. Review.
-
Investigation into the Antigenic Properties and Contributions to Growth in Blood of the Meningococcal Haemoglobin Receptors, HpuAB and HmbR.PLoS One. 2015 Jul 24;10(7):e0133855. doi: 10.1371/journal.pone.0133855. eCollection 2015. PLoS One. 2015. PMID: 26208277 Free PMC article.
-
Sortase independent and dependent systems for acquisition of haem and haemoglobin in Listeria monocytogenes.Mol Microbiol. 2011 Jun;80(6):1581-97. doi: 10.1111/j.1365-2958.2011.07667.x. Epub 2011 May 6. Mol Microbiol. 2011. PMID: 21545655 Free PMC article.
References
-
- Alifano, P., F. Rivellini, D. Limauro, C. B. Bruni, and M. S. Carlomagno. 1991. A consensus motif common to all Rho-dependent prokaryotic transcription terminators. Cell 64:553-563. - PubMed
-
- Al-Tawfiq, J. A., K. R. Fortney, B. P. Katz, A. F. Hood, C. Elkins, and S. M. Spinola. 2000. An isogenic hemoglobin receptor-deficient mutant of Haemophilus ducreyi is attenuated in the human model of experimental infection. J. Infect. Dis. 181:1049-1054. - PubMed
-
- Angerer, A., S. Enz, M. Ochs, and V. Braun. 1995. Transcriptional regulation of ferric citrate transport in Escherichia coli K-12. Fecl belongs to a new subfamily of sigma 70-type factors that respond to extracytoplasmic stimuli. Mol. Microbiol. 18:163-174. - PubMed
-
- Buisine, N., C. M. Tang, and R. Chalmers. 2002. Transposon-like Correia elements: structure, distribution and genetic exchange between pathogenic Neisseria sp. FEBS Lett. 522:52-58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
