

Drosophila Raf's N terminus contains a novel conserved region and can contribute to torso RTK signaling
- PMID: 20008569
- PMCID: PMC2845340
- DOI: 10.1534/genetics.109.111344
Drosophila Raf's N terminus contains a novel conserved region and can contribute to torso RTK signaling
Abstract
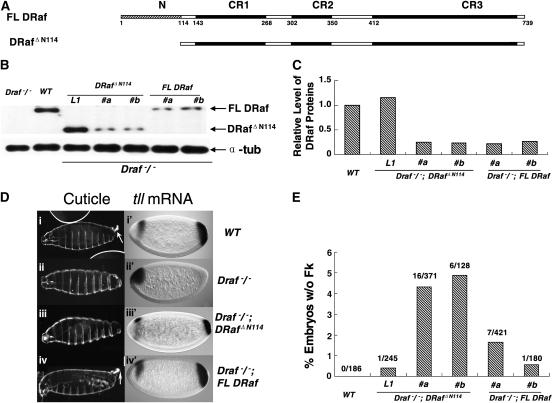
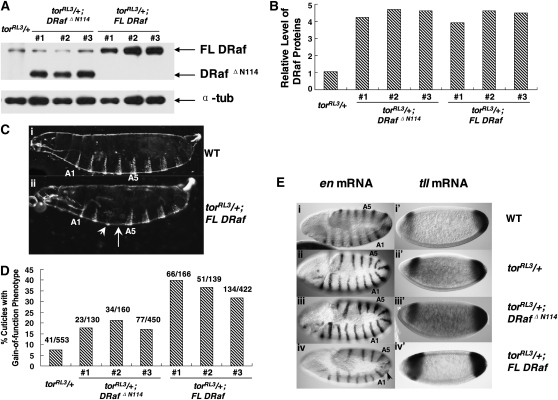
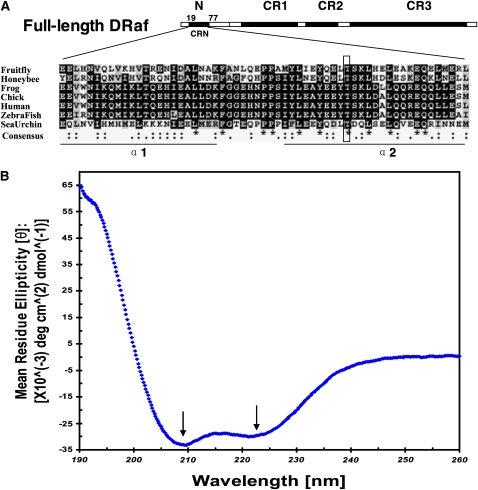
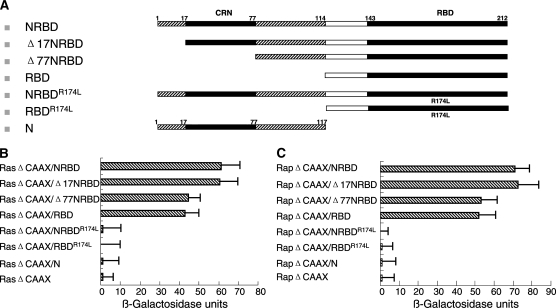
Drosophila Raf (DRaf) contains an extended N terminus, in addition to three conserved regions (CR1-CR3); however, the function(s) of this N-terminal segment remains elusive. In this article, a novel region within Draf's N terminus that is conserved in BRaf proteins of vertebrates was identified and termed conserved region N-terminal (CRN). We show that the N-terminal segment can play a positive role(s) in the Torso receptor tyrosine kinase pathway in vivo, and its contribution to signaling appears to be dependent on the activity of Torso receptor, suggesting this N-terminal segment can function in signal transmission. Circular dichroism analysis indicates that DRaf's N terminus (amino acids 1-117) including CRN (amino acids 19-77) is folded in vitro and has a high content of helical secondary structure as predicted by proteomics tools. In yeast two-hybrid assays, stronger interactions between DRaf's Ras binding domain (RBD) and the small GTPase Ras1, as well as Rap1, were observed when CRN and RBD sequences were linked. Together, our studies suggest that DRaf's extended N terminus may assist in its association with the upstream activators (Ras1 and Rap1) through a CRN-mediated mechanism(s) in vivo.
Figures


Similar articles
-
Ras and Rap: are former enemies now friends?Dev Cell. 2005 Mar;8(3):303-4. doi: 10.1016/j.devcel.2005.02.006. Dev Cell. 2005. PMID: 15737926
-
Ras-independent activation of ERK signaling via the torso receptor tyrosine kinase is mediated by Rap1.Curr Biol. 2005 Feb 22;15(4):366-70. doi: 10.1016/j.cub.2005.02.022. Curr Biol. 2005. PMID: 15723799
-
Apical accumulation of the Sevenless receptor tyrosine kinase during Drosophila eye development is promoted by the small GTPase Rap1.Genetics. 2014 Aug;197(4):1237-50. doi: 10.1534/genetics.114.166272. Epub 2014 Jun 3. Genetics. 2014. PMID: 24899161 Free PMC article.
-
Functions and mechanisms of receptor tyrosine kinase Torso signaling: lessons from Drosophila embryonic terminal development.Dev Dyn. 2005 Mar;232(3):656-72. doi: 10.1002/dvdy.20295. Dev Dyn. 2005. PMID: 15704136 Free PMC article. Review.
-
Ras assemblies and signaling at the membrane.Curr Opin Struct Biol. 2020 Jun;62:140-148. doi: 10.1016/j.sbi.2020.01.009. Epub 2020 Feb 4. Curr Opin Struct Biol. 2020. PMID: 32032914 Review.
Cited by
-
Targeting Aberrant RAS/RAF/MEK/ERK Signaling for Cancer Therapy.Cells. 2020 Jan 13;9(1):198. doi: 10.3390/cells9010198. Cells. 2020. PMID: 31941155 Free PMC article. Review.
-
Similar but different: distinct roles for KRAS and BRAF oncogenes in colorectal cancer development and therapy resistance.Oncotarget. 2015 Aug 28;6(25):20785-800. doi: 10.18632/oncotarget.4750. Oncotarget. 2015. PMID: 26299805 Free PMC article. Review.
-
Mechanistic principles of RAF kinase signaling.Cell Mol Life Sci. 2011 Feb;68(4):553-65. doi: 10.1007/s00018-010-0520-6. Epub 2010 Sep 6. Cell Mol Life Sci. 2011. PMID: 20820846 Free PMC article. Review.
References
-
- Baek, K.-H., J. R. Fabian, F. Sprenger, D. K. Morrison and L. Ambrosio, 1996. The activity of D-raf in torso signal transduction is altered by serine substitution, N terminal deletion and membrane targeting. Dev. Biol. 175 191–204. - PubMed
-
- Blom, N., S. Gammeltoft and S. Brunak, 1999. Sequence- and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 294 1351–1362. - PubMed
-
- Casanova, J., M. Furriols, C. A. Mccormick and G. Struhl, 1995. Similarities between trunk and spätzle, putative extracellular ligands specifying body pattern in Drosophila. Genes Dev. 9 2539–2544. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
