

Human POT1 is required for efficient telomere C-rich strand replication in the absence of WRN
- PMID: 20008939
- PMCID: PMC2800086
- DOI: 10.1101/gad.544009
Human POT1 is required for efficient telomere C-rich strand replication in the absence of WRN
Abstract
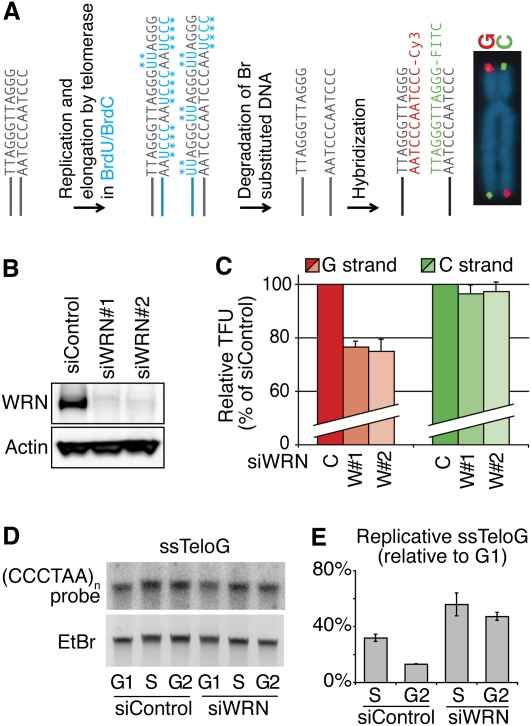
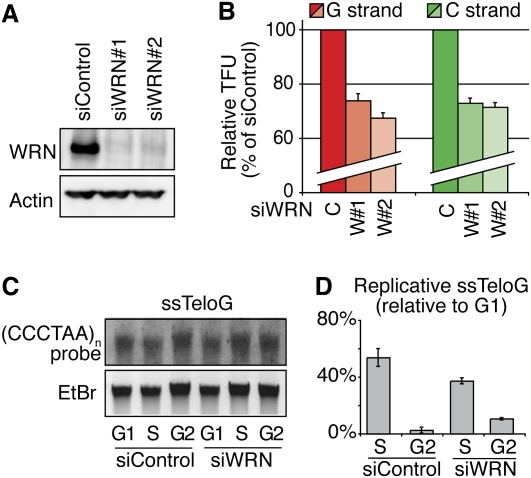
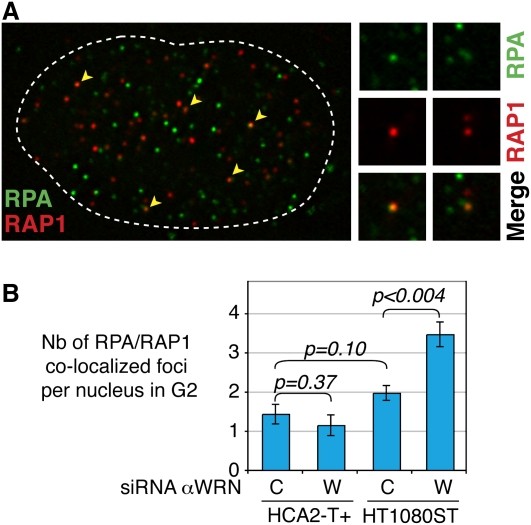
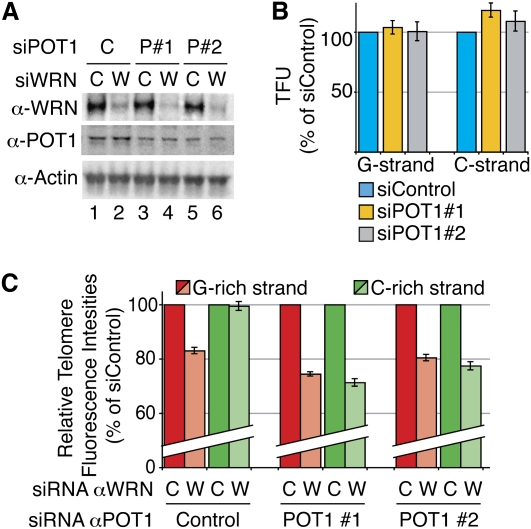
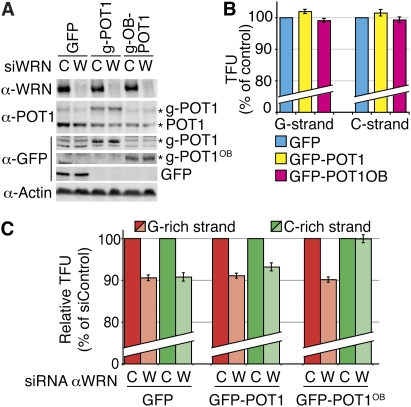
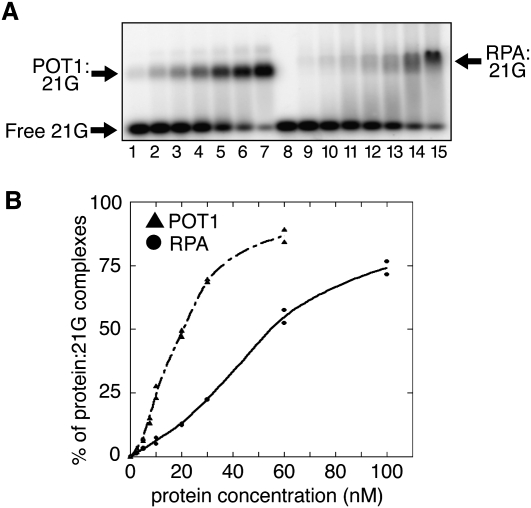
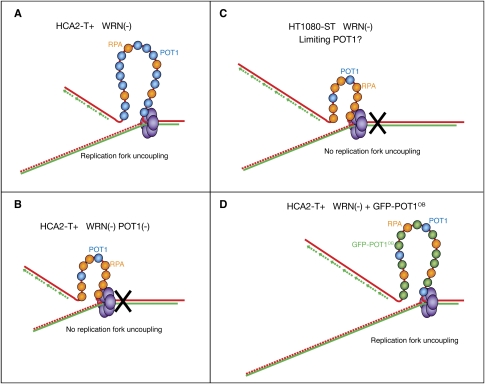
Mechanisms of telomere replication remain poorly defined. It has been suggested that G-rich telomeric strand replication by lagging mechanisms requires, in a stochastic way, the WRN protein. Here we show that this requirement is more systematic than previously thought. Our data are compatible with a situation in which, in the absence of WRN, DNA synthesis at replication forks is uncoupled, thus allowing replication to continue on the C strand, while single G strands accumulate. We also show that in cells in which both WRN and POT1 are limiting, both G- and C-rich telomeric strands shorten, suggesting a complete replication block. Under this particular condition, expression of a fragment spanning the two POT1-OB (oligonucleotide-binding) fold domains is able to restore C (but not G) strand replication, suggesting that binding of POT1 to the lagging strand allows DNA synthesis uncoupling in the absence of WRN. Furthermore, in vitro experiments indicate that purified POT1 has a higher affinity for the telomeric G-rich strand than purified RPA. We propose a model in which the relative enrichments of POT1 versus RPA on the telomeric lagging strand allows or does not allow uncoupling of DNA synthesis at the replication fork. Our study reveals an unanticipated role for hPOT1 during telomere replication.
Figures
References
-
- Alani E, Thresher R, Griffith JD, Kolodner RD. Characterization of DNA-binding and strand-exchange stimulation properties of y-RPA, a yeast single-strand-DNA-binding protein. J Mol Biol. 1992;227:54–71. - PubMed
-
- Arnoult N, Shin-Ya K, Londoño-Vallejo JA. Studying telomere replication by Q-CO-FISH: The effect of telomestatin, a potent G-quadruplex ligand. Cytogenet Genome Res. 2008;122:229–236. - PubMed
-
- Bae SH, Bae KH, Kim JA, Seo YS. RPA governs endonuclease switching during processing of Okazaki fragments in eukaryotes. Nature. 2001;412:456–461. - PubMed
-
- Baird DM, Davis T, Rowson J, Jones CJ, Kipling D. Normal telomere erosion rates at the single cell level in Werner syndrome fibroblast cells. Hum Mol Genet. 2004;13:1515–1524. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous