

Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and Arabidopsis
- PMID: 20009022
- PMCID: PMC2814508
- DOI: 10.1105/tpc.109.070441
Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and Arabidopsis
Abstract
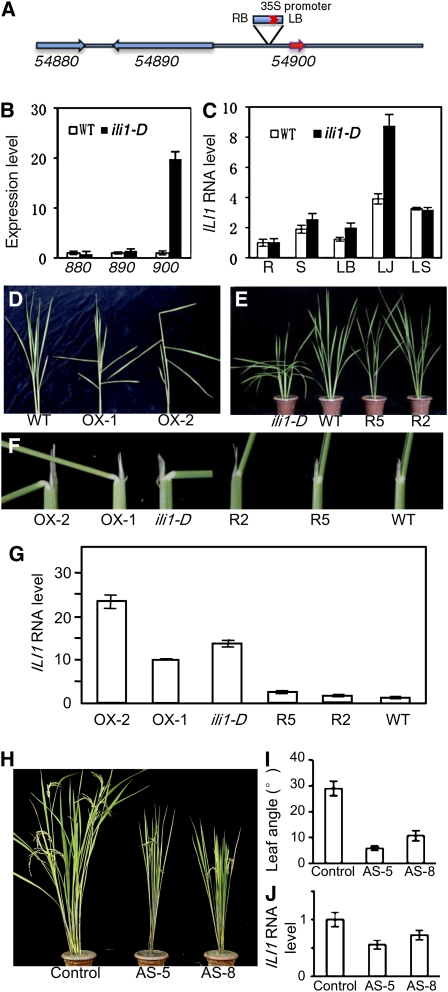
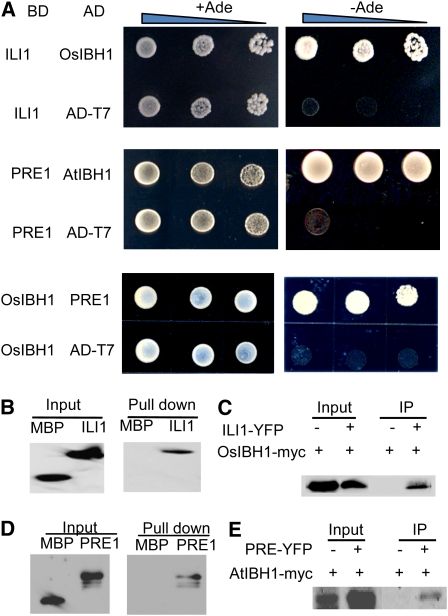
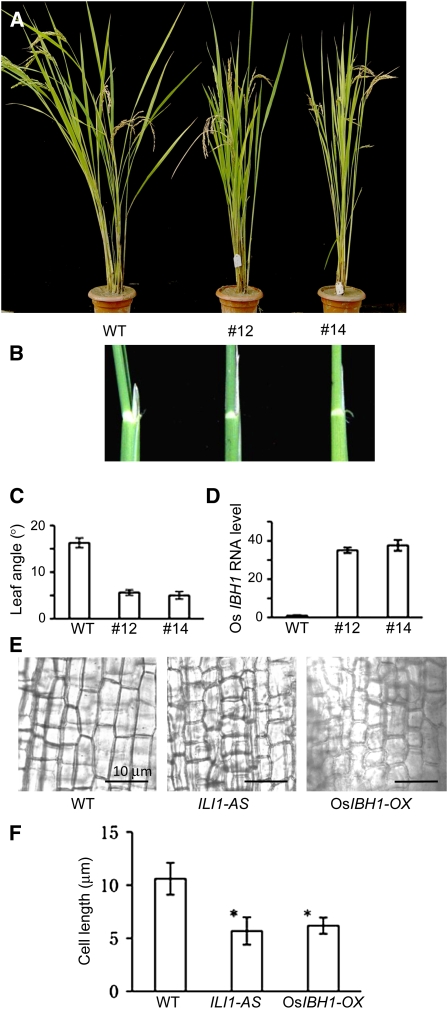
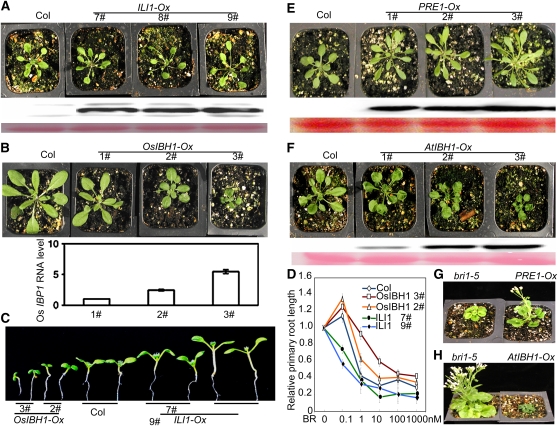
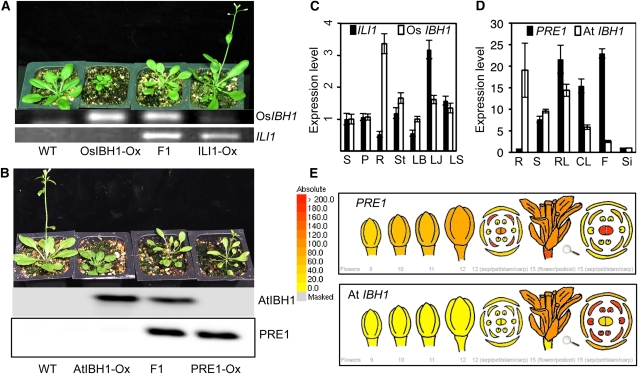
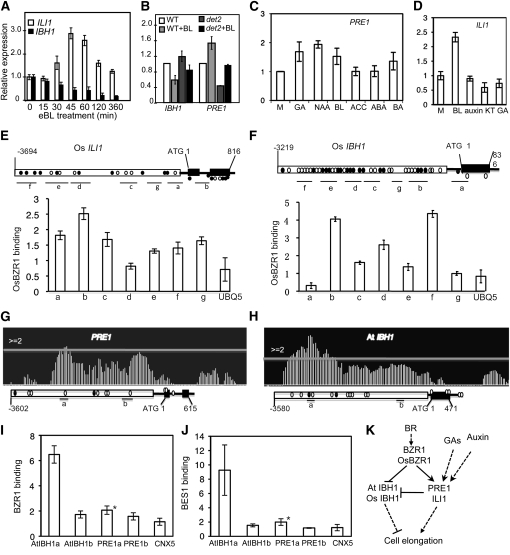
In rice (Oryza sativa), brassinosteroids (BRs) induce cell elongation at the adaxial side of the lamina joint to promote leaf bending. We identified a rice mutant (ili1-D) showing an increased lamina inclination phenotype similar to that caused by BR treatment. The ili1-D mutant overexpresses an HLH protein homologous to Arabidopsis thaliana Paclobutrazol Resistance1 (PRE1) and the human Inhibitor of DNA binding proteins. Overexpression and RNA interference suppression of ILI1 increase and reduce, respectively, rice laminar inclination, confirming a positive role of ILI1 in leaf bending. ILI1 and PRE1 interact with basic helix-loop-helix (bHLH) protein IBH1 (ILI1 binding bHLH), whose overexpression causes erect leaf in rice and dwarfism in Arabidopsis. Overexpression of ILI1 or PRE1 increases cell elongation and suppresses dwarf phenotypes caused by overexpression of IBH1 in Arabidopsis. Thus, ILI1 and PRE1 may inactivate inhibitory bHLH transcription factors through heterodimerization. BR increases the RNA levels of ILI1 and PRE1 but represses IBH1 through the transcription factor BZR1. The spatial and temporal expression patterns support roles of ILI1 in laminar joint bending and PRE1/At IBH1 in the transition from growth of young organs to growth arrest. These results demonstrate a conserved mechanism of BR regulation of plant development through a pair of antagonizing HLH/bHLH transcription factors that act downstream of BZR1 in Arabidopsis and rice.
Figures

References
-
- Bernhardt, C., Zhao, M., Gonzalez, A., Lloyd, A., and Schiefelbein, J. (2005). The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development 132 291–298. - PubMed
-
- Chen, K.Y., Cong, B., Wing, R., Vrebalov, J., and Tanksley, S.D. (2007). Changes in regulation of a transcription factor lead to autogamy in cultivated tomatoes. Science 318 643–645. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
