

Sustained activity of the EGF receptor is an absolute requisite for LH-induced oocyte maturation and cumulus expansion
- PMID: 20009084
- PMCID: PMC5428124
- DOI: 10.1210/me.2009-0267
Sustained activity of the EGF receptor is an absolute requisite for LH-induced oocyte maturation and cumulus expansion
Abstract
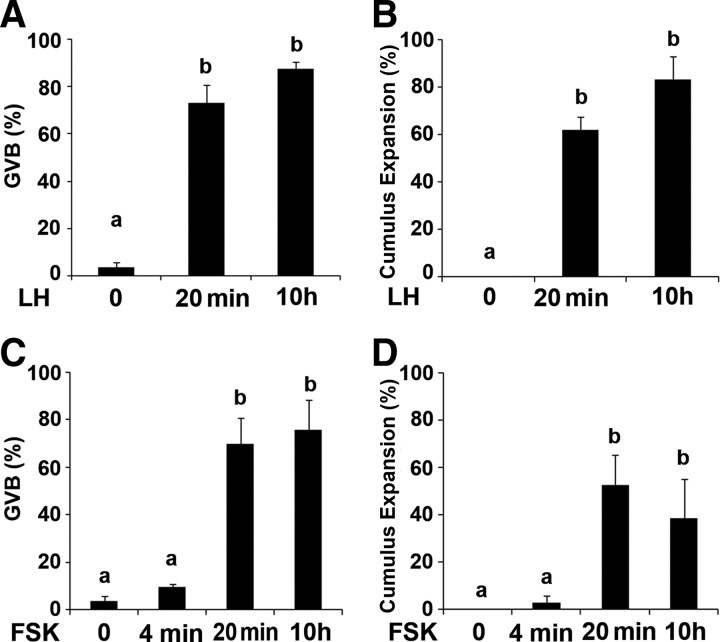
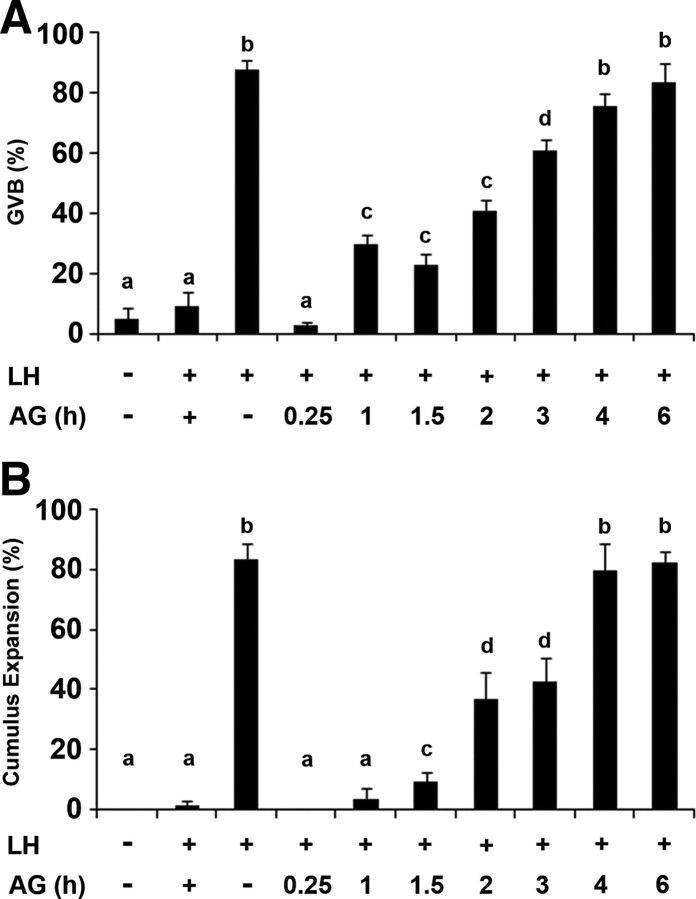
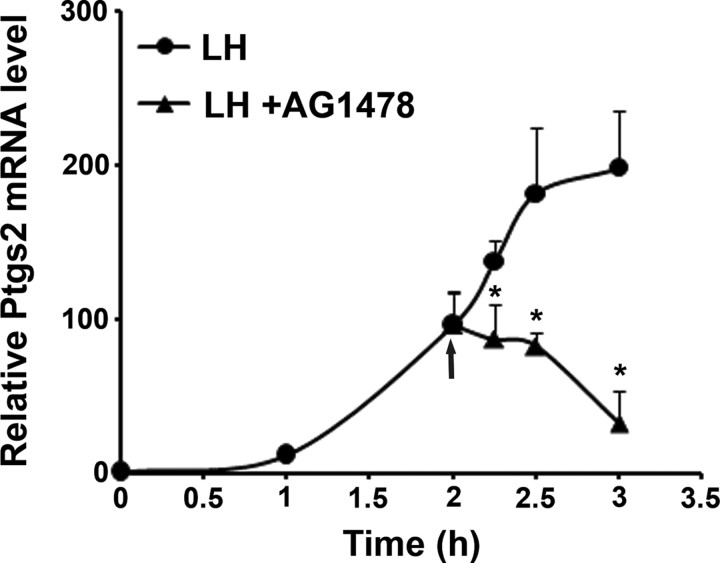
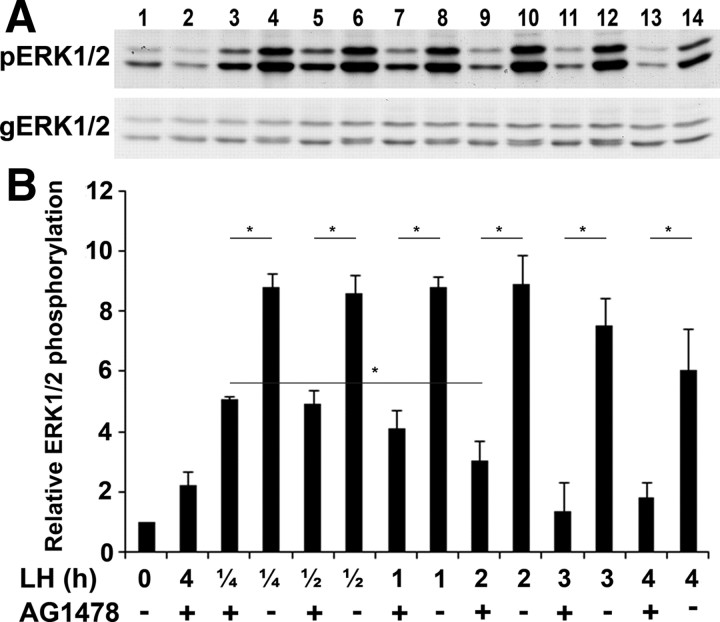
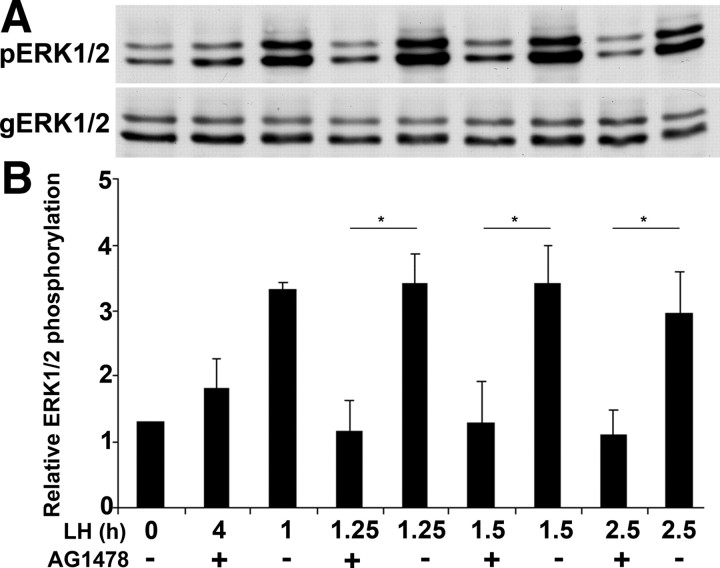
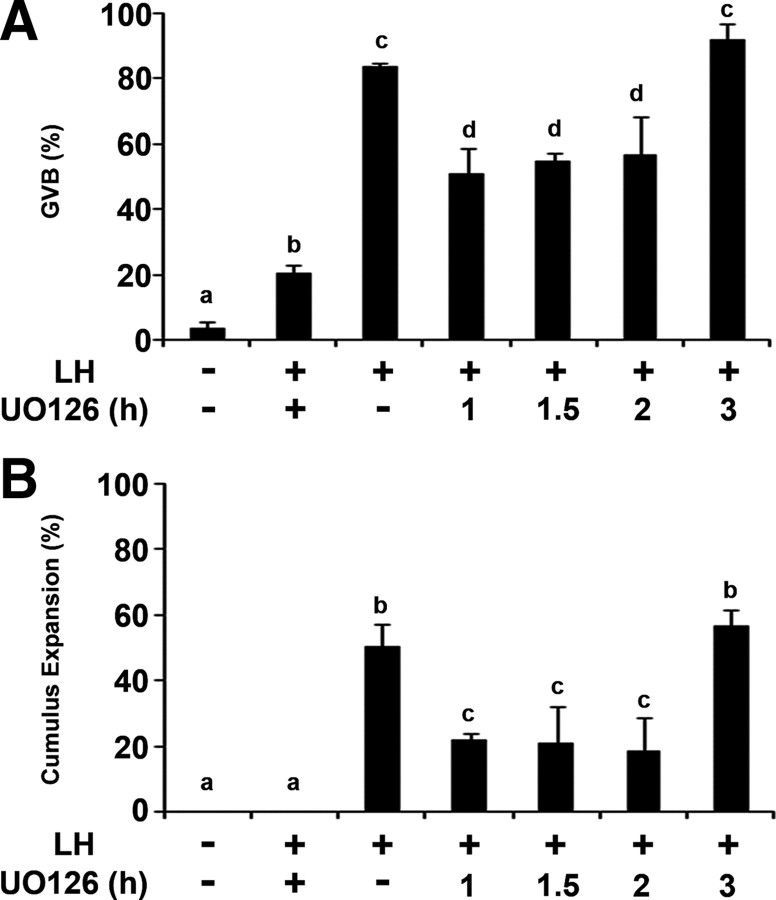
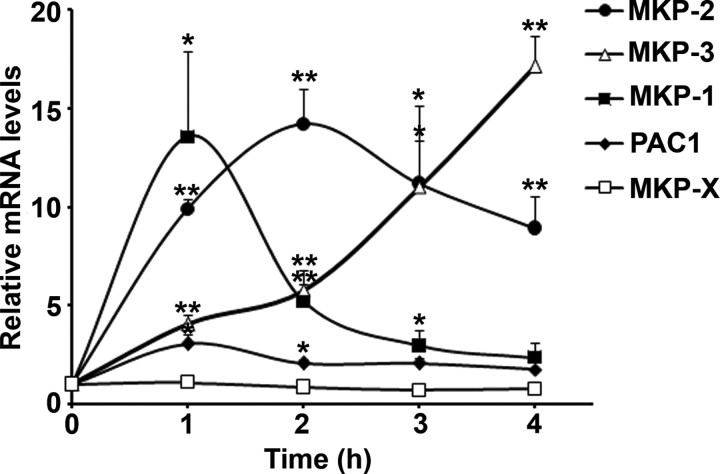
Mammalian reproduction depends on the release of a mature oocyte from the ovarian follicle. Maturation of the oocyte and rupture of the follicle wall constitute part of the responses to the preovulatory surge of LH, which also include cumulus expansion and granulosa cell luteinization. It was previously shown that the epidermal growth factor receptor (EGFR) mediates the ovulatory response to LH in the ovarian follicle. We hypothesized that it is a sustained activity of the EGFR that generates oocyte maturation and cumulus expansion. We demonstrated that, whereas a transient exposure of rat isolated, intact, preovulatory follicles to either LH or forskolin was sufficient to induce oocyte maturation and cumulus expansion, these LH-induced responses were only generated upon a prolonged activity of the EGFR. In addition, the continuous activity of the EGFR is essential for the chronic phosphorylation of the ERK1/2 downstream signaling molecules, which were shown to be essential for oocyte maturation and cumulus expansion. Interestingly, EGFR-sustained activity was also necessary to maintain the up-regulation of Ptgs2, a gene essential for cumulus expansion. The unusual prolonged duration of ERK1/2 activity may possibly be attributed to the late induction of the ERK-specific phosphatase 3, demonstrated herein. These new data shed light on the unique characteristics of EGFR-ERK1/2 activity in the ovarian follicle and emphasize the fact that the ovulatory process involves a nonclassical activation of this pathway.
Figures
Similar articles
-
Synergistic effect between LH and estrogen in the acceleration of cumulus expansion via GPR30 and EGFR pathways.Aging (Albany NY). 2020 Oct 28;12(20):20801-20816. doi: 10.18632/aging.104029. Epub 2020 Oct 28. Aging (Albany NY). 2020. PMID: 33113510 Free PMC article.
-
Hormone-induced expression of tumor necrosis factor alpha-converting enzyme/A disintegrin and metalloprotease-17 impacts porcine cumulus cell oocyte complex expansion and meiotic maturation via ligand activation of the epidermal growth factor receptor.Endocrinology. 2007 Dec;148(12):6164-75. doi: 10.1210/en.2007-0195. Epub 2007 Sep 27. Endocrinology. 2007. PMID: 17901238
-
Mouse oocytes enable LH-induced maturation of the cumulus-oocyte complex via promoting EGF receptor-dependent signaling.Mol Endocrinol. 2010 Jun;24(6):1230-9. doi: 10.1210/me.2009-0497. Epub 2010 Apr 9. Mol Endocrinol. 2010. PMID: 20382892 Free PMC article.
-
The release of EGF domain from EGF-like factors by a specific cleavage enzyme activates the EGFR-MAPK3/1 pathway in both granulosa cells and cumulus cells during the ovulation process.J Reprod Dev. 2012;58(5):510-4. doi: 10.1262/jrd.2012-056. J Reprod Dev. 2012. PMID: 23124701 Review.
-
The epidermal growth factor network: role in oocyte growth, maturation and developmental competence.Hum Reprod Update. 2018 Jan 1;24(1):1-14. doi: 10.1093/humupd/dmx029. Hum Reprod Update. 2018. PMID: 29029246 Review.
Cited by
-
Amphiregulin promotes the maturation of oocytes isolated from the small antral follicles of the rhesus macaque.Hum Reprod. 2012 Aug;27(8):2430-7. doi: 10.1093/humrep/des158. Epub 2012 May 16. Hum Reprod. 2012. PMID: 22593432 Free PMC article.
-
Aberrant activation of KRAS in mouse theca-interstitial cells results in female infertility.Front Physiol. 2022 Aug 19;13:991719. doi: 10.3389/fphys.2022.991719. eCollection 2022. Front Physiol. 2022. PMID: 36060690 Free PMC article.
-
The molecular regulatory mechanisms of meiotic arrest and resumption in Oocyte development and maturation.Reprod Biol Endocrinol. 2023 Oct 2;21(1):90. doi: 10.1186/s12958-023-01143-0. Reprod Biol Endocrinol. 2023. PMID: 37784186 Free PMC article. Review.
-
MAPK3/1 is conducive to luteinizing hormone-mediated C-type natriuretic peptide decrease in bovine granulosa cells.J Reprod Dev. 2016 Apr 22;62(2):137-42. doi: 10.1262/jrd.2015-135. Epub 2015 Dec 10. J Reprod Dev. 2016. PMID: 26655567 Free PMC article.
-
Ovarian dendritic cells act as a double-edged pro-ovulatory and anti-inflammatory sword.Mol Endocrinol. 2014 Jul;28(7):1039-54. doi: 10.1210/me.2013-1400. Epub 2014 May 13. Mol Endocrinol. 2014. PMID: 24825398 Free PMC article.
References
-
- Cameron MR, Foster JS, Bukovsky A, Wimalasena J1996. Activation of mitogen-activated protein kinases by gonadotropins and cyclic adenosine 5′-monophosphates in porcine granulosa cells. Biol Reprod 55:111–119 - PubMed
-
- Das S, Maizels ET, DeManno D, St Clair E, Adam SA, Hunzicker-Dunn M1996. A stimulatory role of cyclic adenosine 3′,5′-monophosphate in follicle-stimulating hormone-activated mitogen-activated protein kinase signaling pathway in rat ovarian granulosa cells. Endocrinology 137:967–974 - PubMed
-
- Richards JS, Rolfes AI1980. Hormonal regulation of cyclic AMP binding to specific receptor proteins in rat ovarian follicles. Characterization by photoaffinity labeling. J Biol Chem 255:5481–5489 - PubMed
-
- Richards JS2001. Perspective: the ovarian follicle—a perspective in 2001. Endocrinology 142:2184–2193 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
