

Discovery of candidate disease genes in ENU-induced mouse mutants by large-scale sequencing, including a splice-site mutation in nucleoredoxin
- PMID: 20011118
- PMCID: PMC2782131
- DOI: 10.1371/journal.pgen.1000759
Discovery of candidate disease genes in ENU-induced mouse mutants by large-scale sequencing, including a splice-site mutation in nucleoredoxin
Abstract
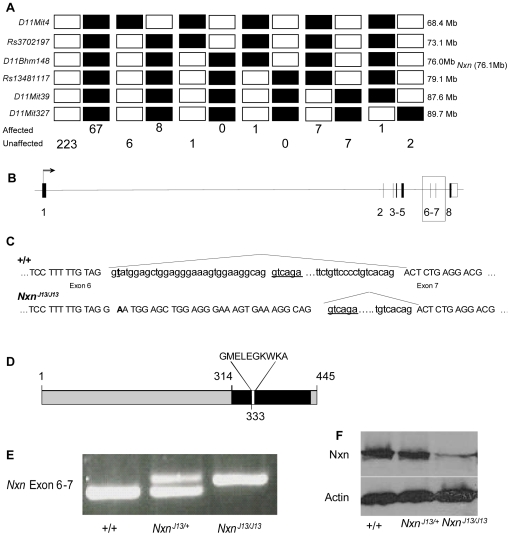
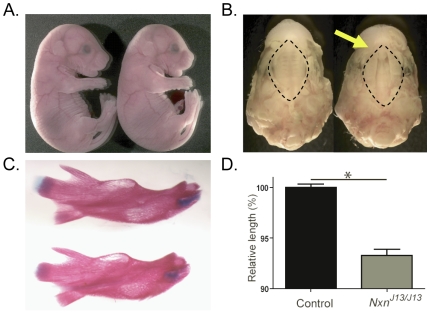
An accurate and precisely annotated genome assembly is a fundamental requirement for functional genomic analysis. Here, the complete DNA sequence and gene annotation of mouse Chromosome 11 was used to test the efficacy of large-scale sequencing for mutation identification. We re-sequenced the 14,000 annotated exons and boundaries from over 900 genes in 41 recessive mutant mouse lines that were isolated in an N-ethyl-N-nitrosourea (ENU) mutation screen targeted to mouse Chromosome 11. Fifty-nine sequence variants were identified in 55 genes from 31 mutant lines. 39% of the lesions lie in coding sequences and create primarily missense mutations. The other 61% lie in noncoding regions, many of them in highly conserved sequences. A lesion in the perinatal lethal line l11Jus13 alters a consensus splice site of nucleoredoxin (Nxn), inserting 10 amino acids into the resulting protein. We conclude that point mutations can be accurately and sensitively recovered by large-scale sequencing, and that conserved noncoding regions should be included for disease mutation identification. Only seven of the candidate genes we report have been previously targeted by mutation in mice or rats, showing that despite ongoing efforts to functionally annotate genes in the mammalian genome, an enormous gap remains between phenotype and function. Our data show that the classical positional mapping approach of disease mutation identification can be extended to large target regions using high-throughput sequencing.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Adams DJ, Quail MA, Cox T, van der Weyden L, Gorick BD, et al. A genome-wide, end-sequenced 129Sv BAC library resource for targeting vector construction. Genomics. 2005;86:753–758. - PubMed
-
- Waterston RH, Lindblad-Toh K, Birney E, Rogers J, Abril JF, et al. Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. - PubMed
-
- Pershouse M, Li J, Yang C, Su H, Brundage E, et al. BAC contig from a 3-cM region of mouse chromosome 11 surrounding Brca1. Genomics. 2000;69:139–142. - PubMed
-
- Zheng B, Sage M, Cai WW, Thompson DM, Tavsanli BC, et al. Engineering a balancer chromosome in the mouse. Nature Genetics. 1999b;22:375–378. - PubMed
-
- Kile BT, Hentges KE, Clark AT, Nakamura H, Salinger AP, et al. Functional genetic analysis of mouse chromosome 11. Nature. 2003;425:81–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
