

Cloning, expression and purification of an acetoacetyl CoA thiolase from sunflower cotyledon
- PMID: 20011134
- PMCID: PMC2793306
- DOI: 10.7150/ijbs.5.736
Cloning, expression and purification of an acetoacetyl CoA thiolase from sunflower cotyledon
Abstract
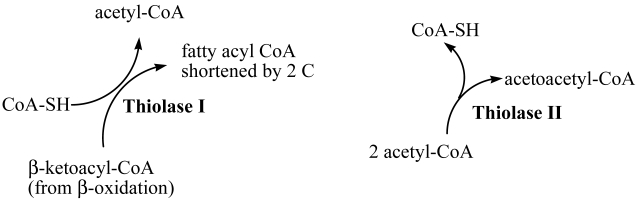
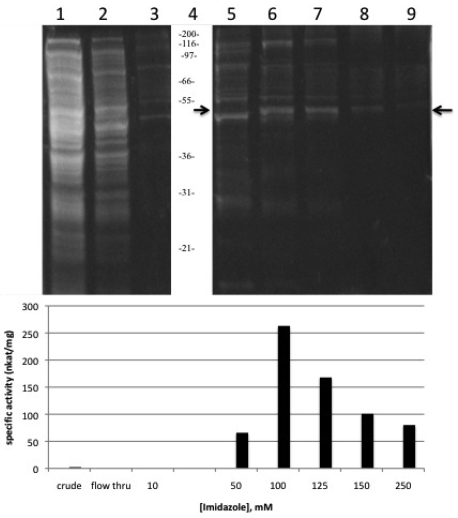
Thiolase I and II coexist as part of the glyoxysomal beta-oxidation system in sunflower (Helianthus annuus L.) cotyledons, the only system shown to have both forms. The importance of thiolases can be underscored not only by their ubiquity, but also by their involvement in a wide variety of processes in plants, animals and bacteria. Here we describe the cloning, expression and purification of acetoacetyl CoA thiolase (AACT) in enzymatically active form. Use of the extensive amount of sequence information from the databases facilitated the efficient generation of the gene-specific primers used in the RACE protocols. The recombinant AACT (1233 bp) shares 75% similarity with other plant AACTs. Comparison of specific activity of this recombinant AACT to a previously reported enzyme purified from primary sunflower cotyledon tissue was very similar (263 nkat/mg protein vs 220 nkat/mg protein, respectively). Combining the most pure fractions from the affinity column, the enzyme was purified 88-fold with a 55% yield of the enzymatically active, 47 kDa AACT.
Keywords: acetoacetyl CoA; cloning; expression; sunflower; thiolase.
Conflict of interest statement
Conflict of Interest: The authors have declared that no conflict of interest exists.
Figures



Similar articles
-
Cloning, expression, and purification of glyoxysomal 3-oxoacyl-CoA thiolase from sunflower cotyledons.Protein Expr Purif. 2004 Jan;33(1):25-33. doi: 10.1016/j.pep.2003.08.022. Protein Expr Purif. 2004. PMID: 14680958
-
Glyoxysomal acetoacetyl-CoA thiolase and 3-oxoacyl-CoA thiolase from sunflower cotyledons.Planta. 2002 Feb;214(4):597-607. doi: 10.1007/s004250100645. Planta. 2002. PMID: 11925043
-
Expression, identification and purification of Dictyostelium acetoacetyl-coa thiolase expressed in Escherichia coli.Int J Biol Sci. 2010 Dec 30;7(1):9-17. doi: 10.7150/ijbs.7.9. Int J Biol Sci. 2010. PMID: 21209787 Free PMC article.
-
Molecular cloning and nucleotide sequence of complementary DNA for human hepatic cytosolic acetoacetyl-coenzyme A thiolase.Biochem Biophys Res Commun. 1994 May 30;201(1):478-85. doi: 10.1006/bbrc.1994.1726. Biochem Biophys Res Commun. 1994. PMID: 7911016
-
Understanding the function of bacterial and eukaryotic thiolases II by integrating evolutionary and functional approaches.Gene. 2014 Jan 1;533(1):5-10. doi: 10.1016/j.gene.2013.09.096. Epub 2013 Oct 11. Gene. 2014. PMID: 24120621 Review.
Cited by
-
Exploration of genes encoding KEGG pathway enzymes in rhizospheric microbiome of the wild plant Abutilon fruticosum.AMB Express. 2024 Feb 21;14(1):27. doi: 10.1186/s13568-024-01678-4. AMB Express. 2024. PMID: 38381255 Free PMC article.
-
Molecular Cloning, Characterization, and Functional Analysis of Acetyl-CoA C-Acetyltransferase and Mevalonate Kinase Genes Involved in Terpene Trilactone Biosynthesis from Ginkgo biloba.Molecules. 2017 Jan 2;22(1):74. doi: 10.3390/molecules22010074. Molecules. 2017. PMID: 28045448 Free PMC article.
-
The Cytosolic Acetoacetyl-CoA Thiolase TaAACT1 Is Required for Defense against Fusarium pseudograminearum in Wheat.Int J Mol Sci. 2023 Mar 24;24(7):6165. doi: 10.3390/ijms24076165. Int J Mol Sci. 2023. PMID: 37047146 Free PMC article.
-
Salicylic acid-induced upregulation of AtAACT and AtMVD expression enhances triterpene biosynthesis in Athelia termitophila.World J Microbiol Biotechnol. 2025 Feb 27;41(3):87. doi: 10.1007/s11274-025-04301-1. World J Microbiol Biotechnol. 2025. PMID: 40011284
-
Transcriptomics-metabolomics joint analysis: New highlight into the triterpenoid saponin biosynthesis in quinoa (Chenopodium quinoa Willd.).Front Plant Sci. 2022 Oct 19;13:964558. doi: 10.3389/fpls.2022.964558. eCollection 2022. Front Plant Sci. 2022. PMID: 36340365 Free PMC article.
References
-
- Igual JC, González-Bosch C, Dopazo J, Pérez-Ortín JE. Phylogenetic analysis of the thiolase family. Implications for the evolutionary origin of peroxisomes. Journal of Molecular Evolution. 1992;35:147–155. - PubMed
-
- Frevert j, Kindl H. Purification of glyoxysomal acetyl-CoA acyltransferase. Hoppe Seylers Z Physiol Chem. 1980;361:537–542. - PubMed
-
- Carrie C, Murcha MW, Millar AH, Smith SM, Whelan J. Nine 3-ketoacyl-CoA thiolases (KATs) and acetoacetyl-CoA thiolases (ACATs) encoded by five genes in Arabidopsis thaliana are targeted either to peroxisomes or cytosol but not to mitochondria. Plant Molecular Biology. 2007;63:97–108. - PubMed
-
- Masterson C, Wood C. Mitochondrial β-oxidation of fatty acids in higher plants. Physiologia Plantarum. 2000;109:217–224.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
