

Novel functions of photoreceptor guanylate cyclases revealed by targeted deletion
- PMID: 20012162
- PMCID: PMC2832916
- DOI: 10.1007/s11010-009-0322-z
Novel functions of photoreceptor guanylate cyclases revealed by targeted deletion
Abstract
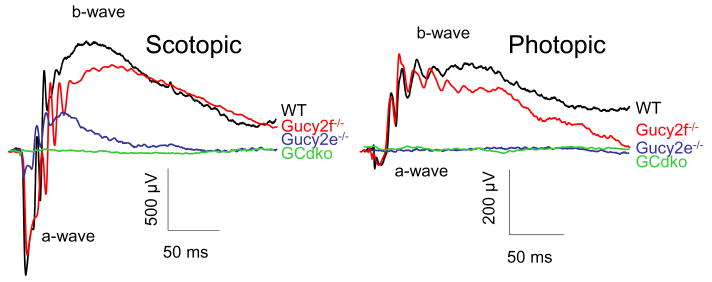
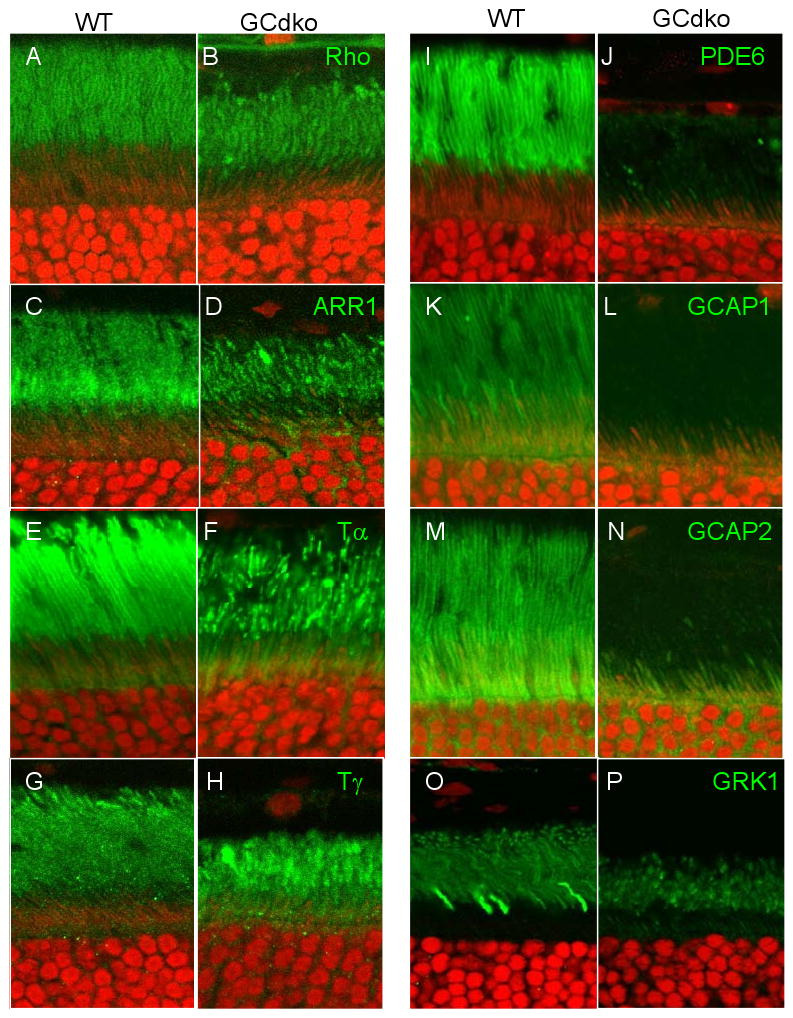
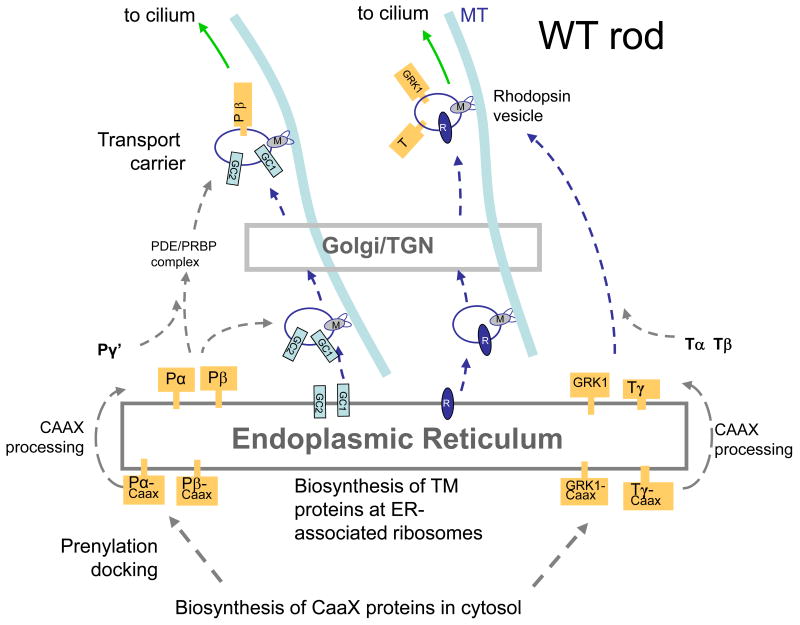
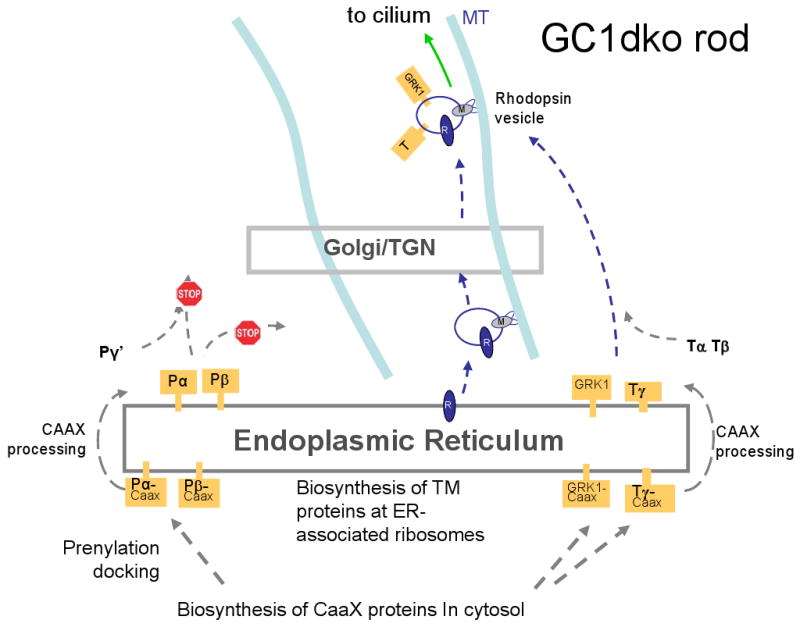
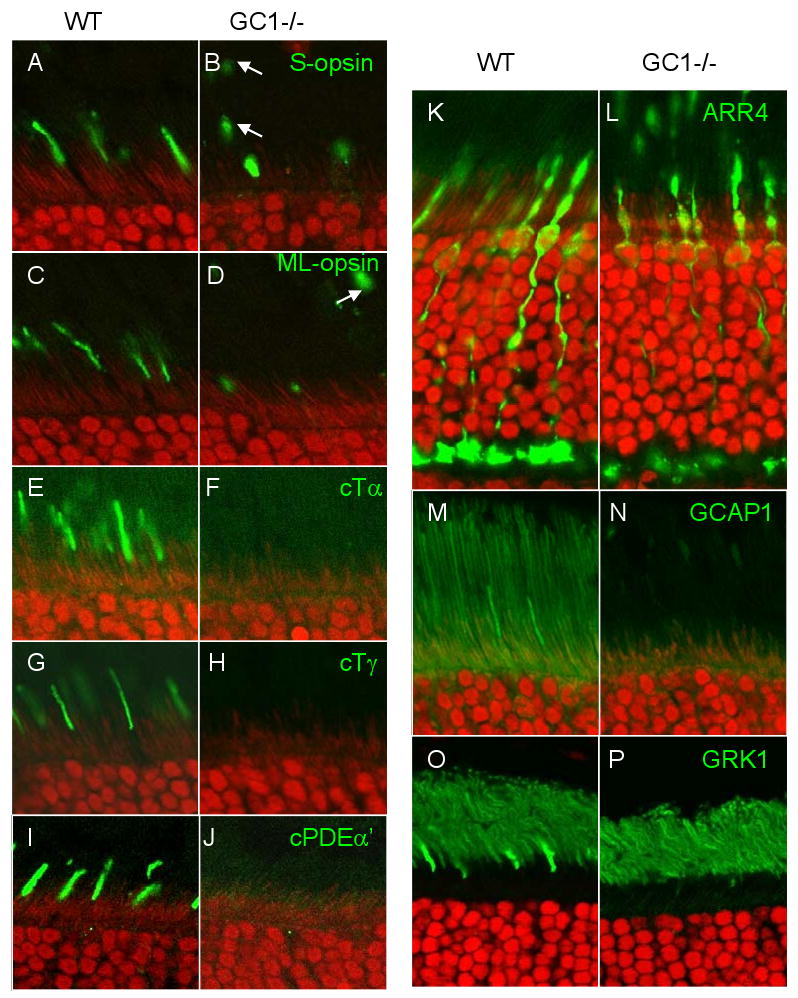
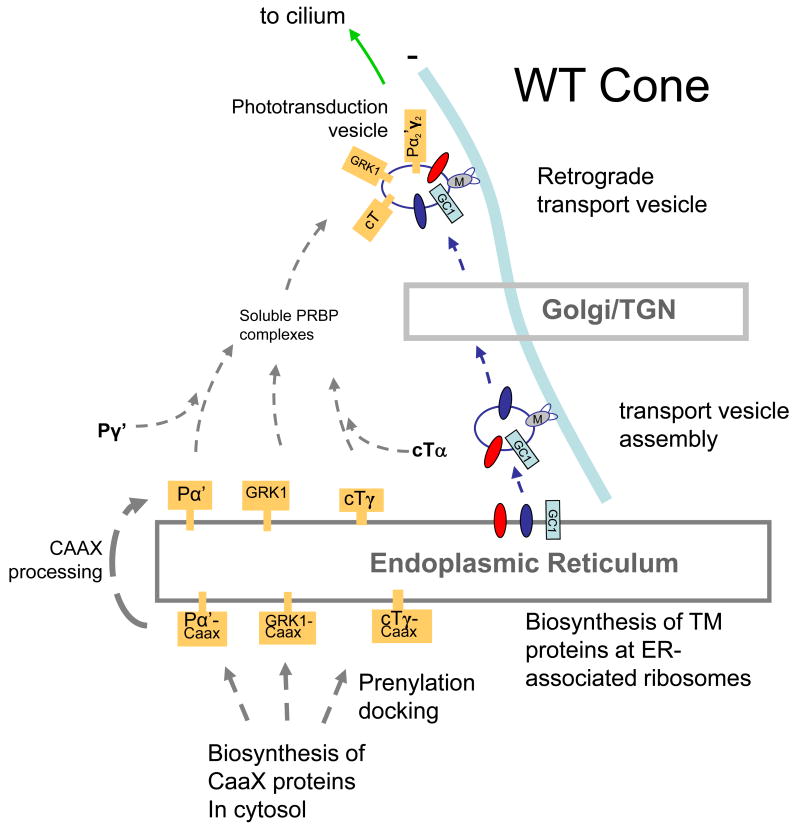
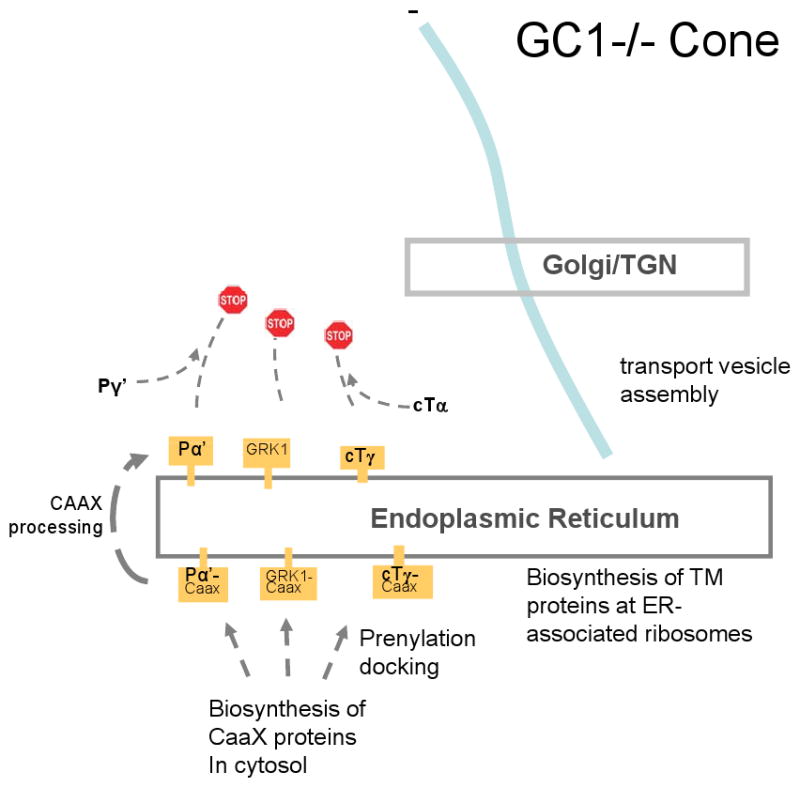
Targeted deletion of membrane guanylate cyclases (GCs) has yielded new information concerning their function. Here, we summarize briefly recent results of laboratory generated non-photoreceptor GC knockouts characterized by complex phenotypes affecting the vasculature, heart, brain, kidney, and other tissues. The main emphasis of the review, however, addresses the two GCs expressed in retinal photoreceptors, termed GC-E and GC-F. Naturally occurring GC-E (GUCY2D) null alleles in human and chicken are associated with an early onset blinding disorder, termed "Leber congenital amaurosis type 1" (LCA-1), characterized by extinguished scotopic and photopic ERGs, and retina degeneration. In mouse, a GC-E null genotype produces a recessive cone dystrophy, while rods remain functional. Rod function is supported by the presence of GC-F (Gucy2f), a close relative of GC-E. Deletion of Gucy2f has very little effect on rod and cone physiology and survival. However, a GC-E/GC-F double knockout (GCdko) phenotypically resembles human LCA-1 with extinguished ERGs and rod/cone degeneration. In GCdko rods, PDE6 and GCAPs are absent in outer segments. In contrast, GC-E(-/-) cones lack proteins of the entire phototransduction cascade. These results suggest that GC-E may participate in transport of peripheral membrane proteins from the endoplasmic reticulum (ER) to the outer segments.
Figures
Similar articles
-
Functional study of two biochemically unusual mutations in GUCY2D Leber congenital amaurosis expressed via adenoassociated virus vector in mouse retinas.Mol Vis. 2016 Nov 10;22:1342-1351. eCollection 2016. Mol Vis. 2016. PMID: 27881908 Free PMC article.
-
Genotype-functional-phenotype correlations in photoreceptor guanylate cyclase (GC-E) encoded by GUCY2D.Prog Retin Eye Res. 2018 Mar;63:69-91. doi: 10.1016/j.preteyeres.2017.10.003. Epub 2017 Oct 20. Prog Retin Eye Res. 2018. PMID: 29061346 Review.
-
GUCY2D Cone-Rod Dystrophy-6 Is a "Phototransduction Disease" Triggered by Abnormal Calcium Feedback on Retinal Membrane Guanylyl Cyclase 1.J Neurosci. 2018 Mar 21;38(12):2990-3000. doi: 10.1523/JNEUROSCI.2985-17.2018. Epub 2018 Feb 12. J Neurosci. 2018. PMID: 29440533 Free PMC article.
-
A model for transport of membrane-associated phototransduction polypeptides in rod and cone photoreceptor inner segments.Vision Res. 2008 Feb;48(3):442-52. doi: 10.1016/j.visres.2007.08.020. Epub 2007 Oct 18. Vision Res. 2008. PMID: 17949773 Free PMC article. Review.
-
The function of guanylate cyclase 1 and guanylate cyclase 2 in rod and cone photoreceptors.J Biol Chem. 2007 Mar 23;282(12):8837-47. doi: 10.1074/jbc.M610369200. Epub 2007 Jan 25. J Biol Chem. 2007. PMID: 17255100 Free PMC article.
Cited by
-
Protein sorting, targeting and trafficking in photoreceptor cells.Prog Retin Eye Res. 2013 Sep;36:24-51. doi: 10.1016/j.preteyeres.2013.03.002. Epub 2013 Apr 3. Prog Retin Eye Res. 2013. PMID: 23562855 Free PMC article. Review.
-
Phosphodiesterase 6C, cGMP-specific cone alpha'.AFCS Nat Mol Pages. 2011;2011:A001756. doi: 10.1038/mp.a001756.01. Epub 2011 Jun 3. AFCS Nat Mol Pages. 2011. PMID: 26504430 Free PMC article.
-
The GAFa domain of phosphodiesterase-6 contains a rod outer segment localization signal.J Neurochem. 2014 Apr;129(2):256-63. doi: 10.1111/jnc.12501. Epub 2013 Nov 20. J Neurochem. 2014. PMID: 24147783 Free PMC article.
-
Control of the Nucleotide Cycle in Photoreceptor Cell Extracts by Retinal Degeneration Protein 3.Front Mol Neurosci. 2018 Feb 21;11:52. doi: 10.3389/fnmol.2018.00052. eCollection 2018. Front Mol Neurosci. 2018. PMID: 29515371 Free PMC article.
-
Insights into the role of RD3 in guanylate cyclase trafficking, photoreceptor degeneration, and Leber congenital amaurosis.Front Mol Neurosci. 2014 May 26;7:44. doi: 10.3389/fnmol.2014.00044. eCollection 2014. Front Mol Neurosci. 2014. PMID: 24904271 Free PMC article. Review.
References
-
- Kuhn M. Structure, regulation, and function of mammalian membrane guanylyl cyclase receptors, with a focus on guanylyl cyclase-A. Circ Res. 2003;93:700–709. - PubMed
-
- Lucas KA, Pitari GM, Kazerounian S, Ruiz-Stewart I, Park J, Schulz S, Chepenik KP, Waldman SA. Guanylyl cyclases and signaling by cyclic GMP. Pharmacol Rev. 2000;52:375–414. - PubMed
-
- Potter LR. Domain analysis of human transmembrane guanylyl cyclase receptors: implications for regulation. Front Biosci. 2005;10:1205–1220. - PubMed
-
- Sitaramayya A. Soluble guanylate cyclases in the retina. Mol Cell Biochem. 2002;230:177–186. - PubMed
-
- Noll GN, Billek M, Pietruck C, Schmidt KF. Inhibition of nitric oxide synthase alters light responses and dark voltage of amphibian photoreceptors. Neuropharmacology. 1994;33:1407–1412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
