

Molecular analysis of the replication program in unicellular model organisms
- PMID: 20012185
- PMCID: PMC3976475
- DOI: 10.1007/s10577-009-9099-x
Molecular analysis of the replication program in unicellular model organisms
Abstract
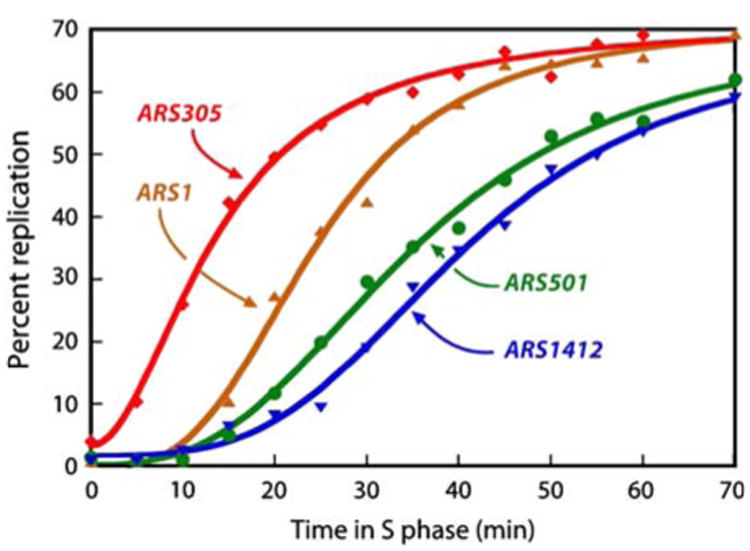
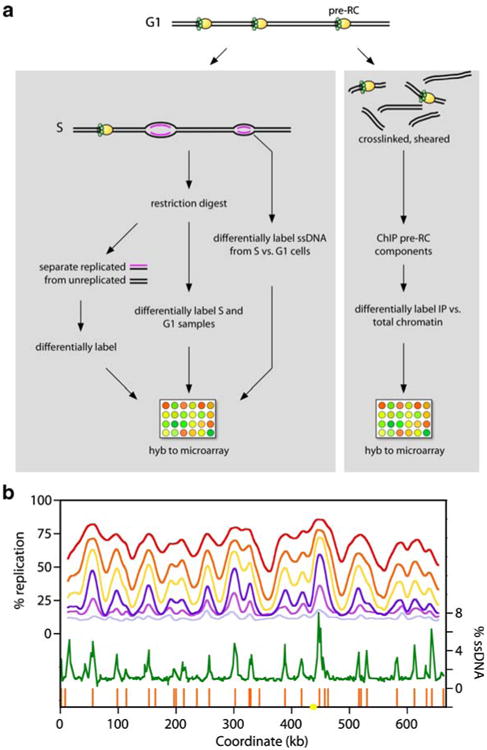
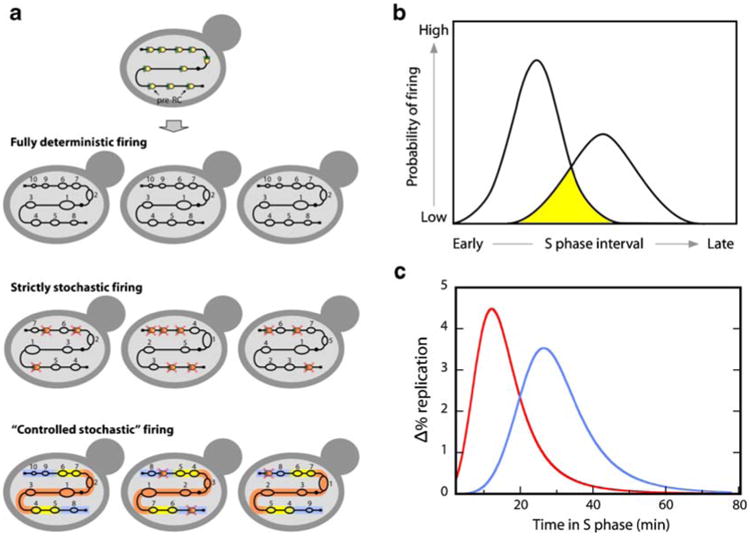
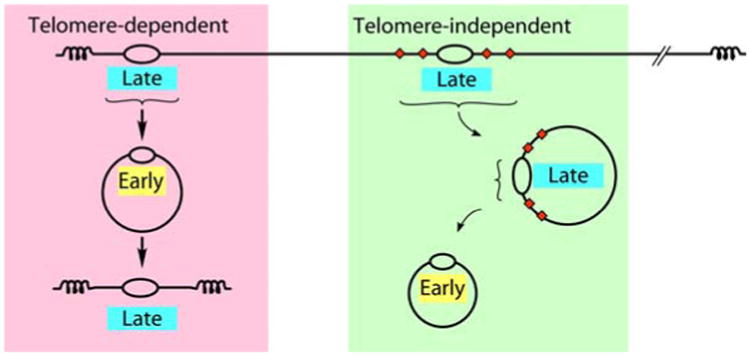
Eukaryotes have long been reported to show temporal programs of replication, different portions of the genome being replicated at different times in S phase, with the added possibility of developmentally regulated changes in this pattern depending on species and cell type. Unicellular model organisms, primarily the budding yeast Saccharomyces cerevisiae, have been central to our current understanding of the mechanisms underlying the regulation of replication origins and the temporal program of replication in particular. But what exactly is a temporal program of replication, and how might it arise? In this article, we explore this question, drawing again on the wealth of experimental information in unicellular model organisms.
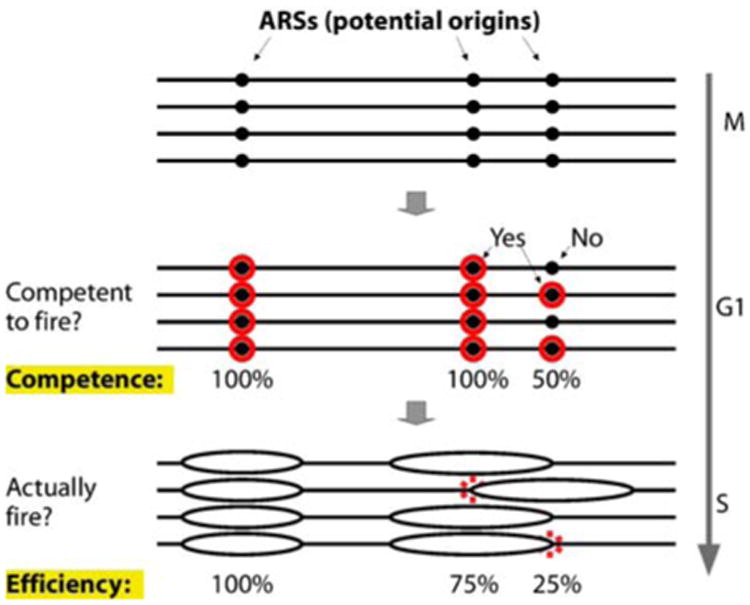
Figures
Similar articles
-
Replication dynamics of the yeast genome.Science. 2001 Oct 5;294(5540):115-21. doi: 10.1126/science.294.5540.115. Science. 2001. PMID: 11588253
-
A model for the spatiotemporal organization of DNA replication in Saccharomyces cerevisiae.Mol Genet Genomics. 2009 Jul;282(1):25-35. doi: 10.1007/s00438-009-0443-9. Epub 2009 Mar 22. Mol Genet Genomics. 2009. PMID: 19306105 Free PMC article.
-
CLB5-dependent activation of late replication origins in S. cerevisiae.Mol Cell. 1998 Aug;2(2):173-82. doi: 10.1016/s1097-2765(00)80127-6. Mol Cell. 1998. PMID: 9734354
-
The dynamics of chromosome replication in yeast.Curr Top Dev Biol. 2003;55:1-73. doi: 10.1016/s0070-2153(03)01001-9. Curr Top Dev Biol. 2003. PMID: 12959193 Review. No abstract available.
-
Replication conrol: choreographing replication origins.Curr Biol. 1998 Oct 22;8(21):R771-3. doi: 10.1016/s0960-9822(07)00483-6. Curr Biol. 1998. PMID: 9799729 Review.
Cited by
-
Replication timing and its emergence from stochastic processes.Trends Genet. 2012 Aug;28(8):374-81. doi: 10.1016/j.tig.2012.03.011. Epub 2012 Apr 18. Trends Genet. 2012. PMID: 22520729 Free PMC article. Review.
-
Mathematical modeling of genome replication.Phys Rev E Stat Nonlin Soft Matter Phys. 2012 Sep;86(3 Pt 1):031916. doi: 10.1103/PhysRevE.86.031916. Epub 2012 Sep 17. Phys Rev E Stat Nonlin Soft Matter Phys. 2012. PMID: 23030953 Free PMC article.
-
Genomic methods for measuring DNA replication dynamics.Chromosome Res. 2020 Mar;28(1):49-67. doi: 10.1007/s10577-019-09624-y. Epub 2019 Dec 17. Chromosome Res. 2020. PMID: 31848781 Free PMC article. Review.
-
The yeast Dbf4 Zn2+ finger domain suppresses single-stranded DNA at replication forks initiated from a subset of origins.Curr Genet. 2022 Apr;68(2):253-265. doi: 10.1007/s00294-022-01230-6. Epub 2022 Feb 11. Curr Genet. 2022. PMID: 35147742 Free PMC article.
-
On the Interplay of the DNA Replication Program and the Intra-S Phase Checkpoint Pathway.Genes (Basel). 2019 Jan 29;10(2):94. doi: 10.3390/genes10020094. Genes (Basel). 2019. PMID: 30700024 Free PMC article. Review.
References
-
- Bell SP, Dutta A. DNA replication in eukaryotic cells. Annu Rev Biochem. 2002;71:333–374. - PubMed
-
- Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature. 1992;357:128–134. - PubMed
-
- Blow JJ, Laskey RA. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature. 1988;332:546–548. - PubMed
-
- Brewer BJ. When polymerases collide: replication and the transcriptional organization of the E. coli chromosome. Cell. 1988;53:679–686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
