

Differential water permeability and regulation of three aquaporin 4 isoforms
- PMID: 20013023
- PMCID: PMC11115813
- DOI: 10.1007/s00018-009-0218-9
Differential water permeability and regulation of three aquaporin 4 isoforms
Abstract
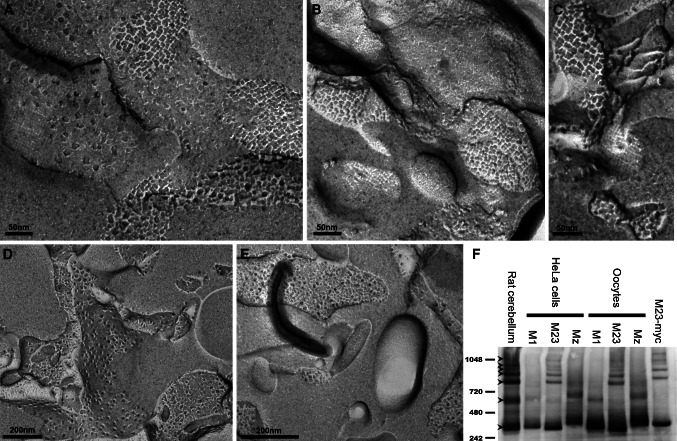
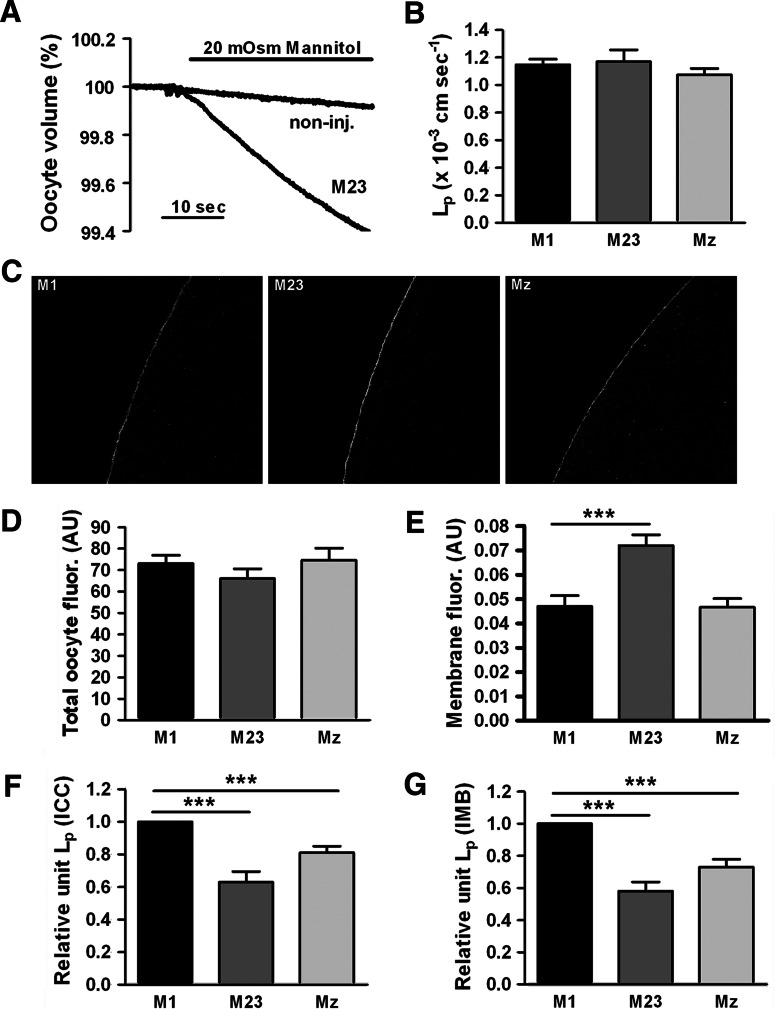
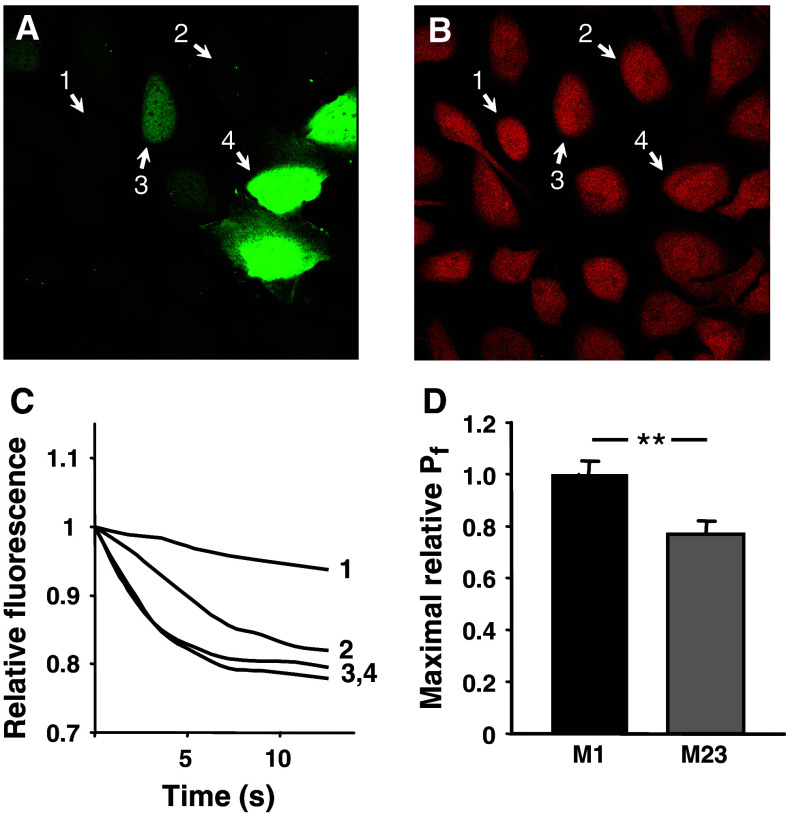
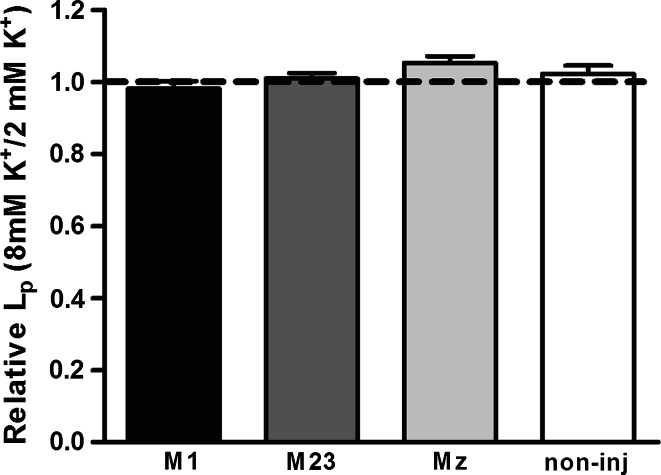
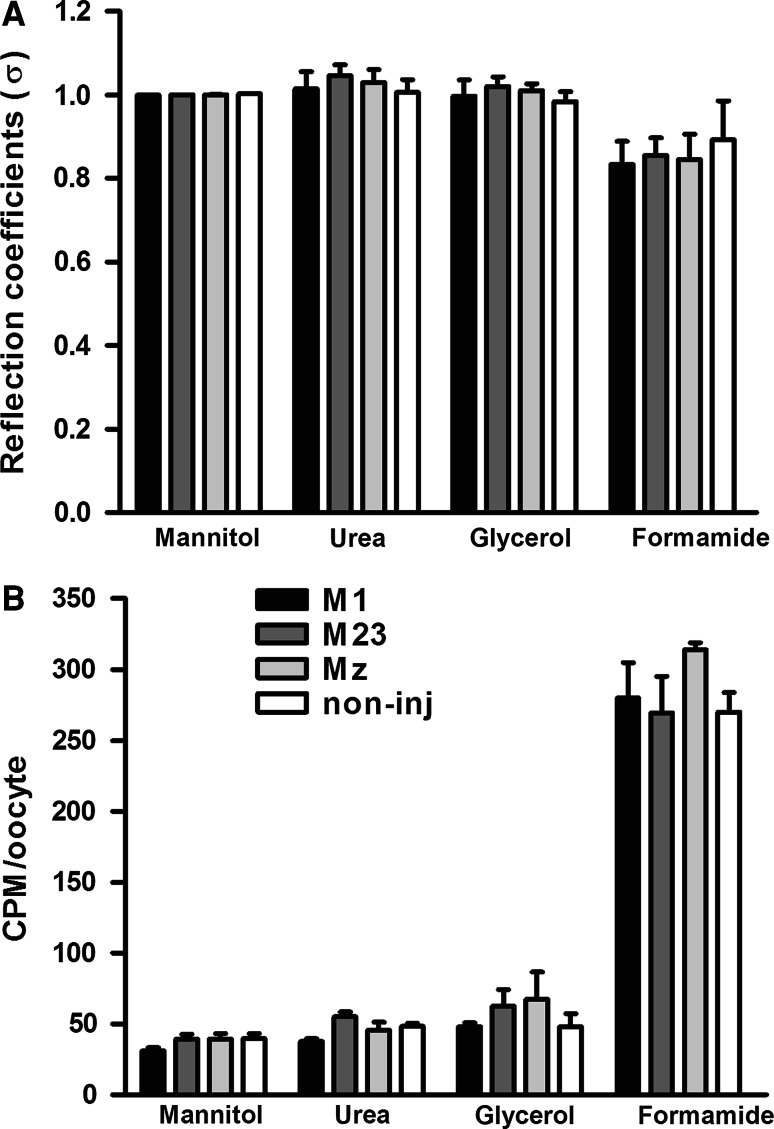
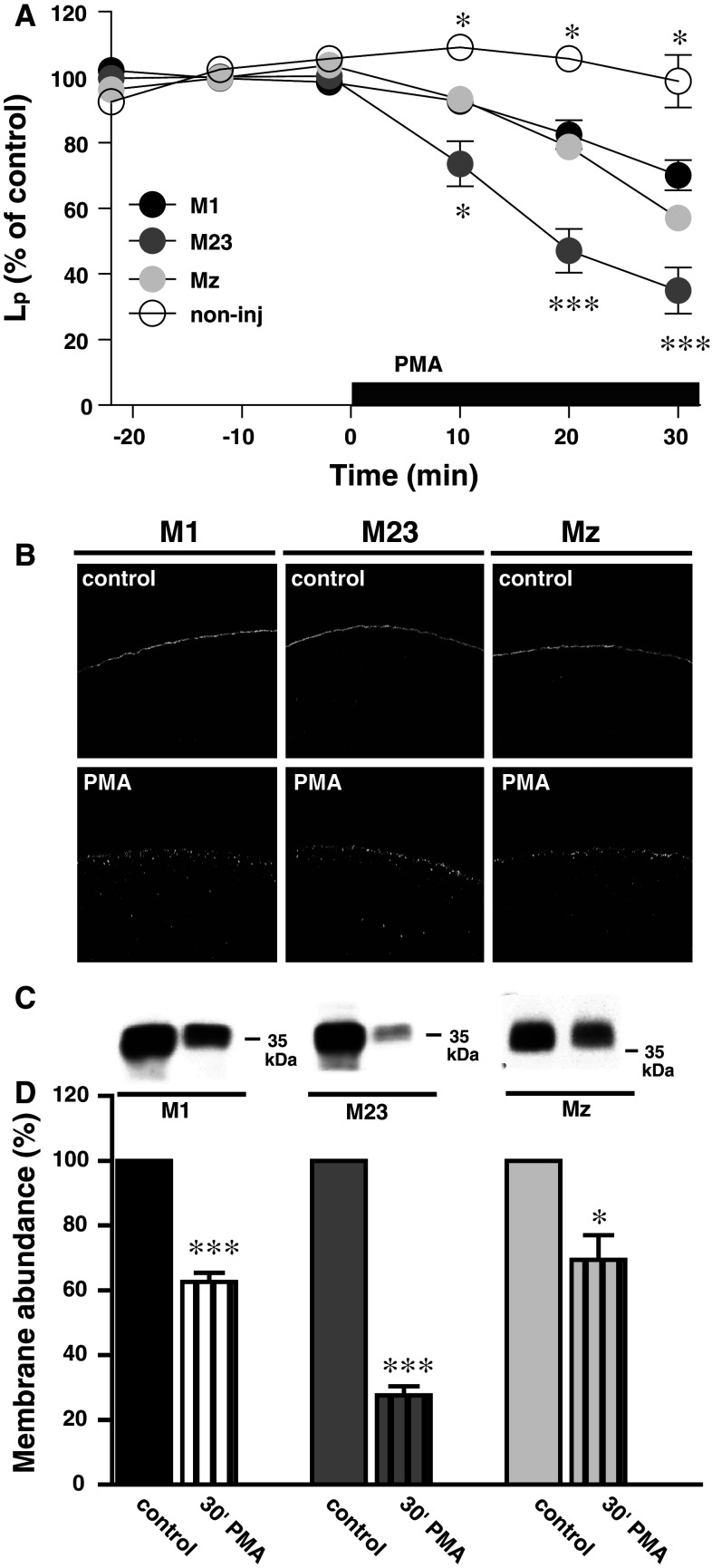
Aquaporin 4 (AQP4) is expressed in the perivascular glial endfeet and is an important pathway for water during formation and resolution of brain edema. In this study, we examined the functional properties and relative unit water permeability of three functional isoforms of AQP4 expressed in the brain (M1, M23, Mz). The M23 isoform gave rise to square arrays when expressed in Xenopus laevis oocytes. The relative unit water permeability differed significantly between the isoforms in the order of M1 > Mz > M23. None of the three isoforms were permeable to small osmolytes nor were they affected by changes in external K(+) concentration. Upon protein kinase C (PKC) activation, oocytes expressing the three isoforms demonstrated rapid reduction of water permeability, which correlated with AQP4 internalization. The M23 isoform was more sensitive to PKC regulation than the longer isoforms and was internalized significantly faster. Our results suggest a specific role for square array formation.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
