

Physiological and glycomic characterization of N-acetylglucosaminyltransferase-IVa and -IVb double deficient mice
- PMID: 20015870
- PMCID: PMC2900882
- DOI: 10.1093/glycob/cwp200
Physiological and glycomic characterization of N-acetylglucosaminyltransferase-IVa and -IVb double deficient mice
Abstract
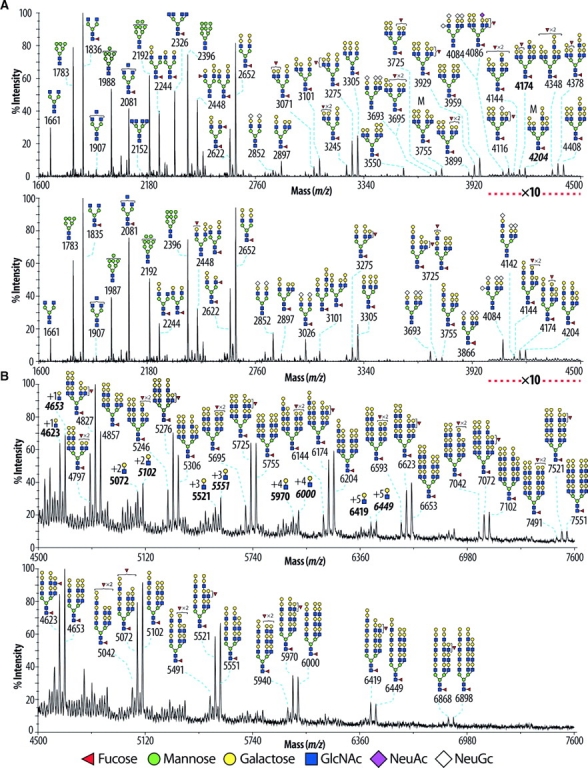
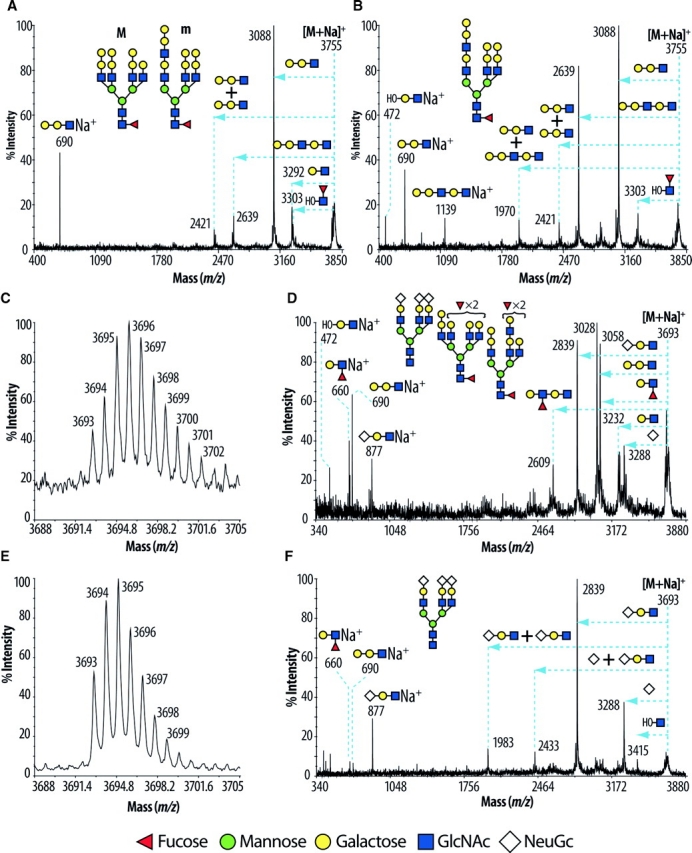
N-Acetylglucosaminyltransferase-IV (GnT-IV) has two isoenzymes, GnT-IVa and GnT-IVb, which initiate the GlcNAcbeta1-4 branch synthesis on the Manalpha1-3 arm of the N-glycan core thereby increasing N-glycan branch complexity and conferring endogenous lectin binding epitopes. To elucidate the physiological significance of GnT-IV, we engineered and characterized GnT-IVb-deficient mice and further generated GnT-IVa/-IVb double deficient mice. In wild-type mice, GnT-IVa expression is restricted to gastrointestinal tissues, whereas GnT-IVb is broadly expressed among organs. GnT-IVb deficiency induced aberrant GnT-IVa expression corresponding to the GnT-IVb distribution pattern that might be attributed to increased Ets-1, which conceivably activates the Mgat4a promoter, and thereafter preserved apparent GnT-IV activity. The compensative GnT-IVa expression might contribute to amelioration of the GnT-IVb-deficient phenotype. GnT-IVb deficiency showed mild phenotypic alterations in hematopoietic cell populations and hemostasis. GnT-IVa/-IVb double deficiency completely abolished GnT-IV activity that resulted in the disappearance of the GlcNAcbeta1-4 branch on the Manalpha1-3 arm that was confirmed by MALDI-TOF MS and GC-MS linkage analyses. Comprehensive glycomic analyses revealed that the abundance of terminal moieties was preserved in GnT-IVa/-IVb double deficiency that was due to the elevated expression of glycosyltransferases regarding synthesis of terminal moieties. Thereby, this may maintain the expression of glycan ligands for endogenous lectins and prevent cellular dysfunctions. The fact that the phenotype of GnT-IVa/-IVb double deficiency largely overlapped that of GnT-IVa single deficiency can be attributed to the induced glycomic compensation. This is the first report that mammalian organs have highly organized glycomic compensation systems to preserve N-glycan branch complexity.
Figures
Similar articles
-
Examination of differential glycoprotein preferences of N-acetylglucosaminyltransferase-IV isozymes a and b.J Biol Chem. 2022 Sep;298(9):102400. doi: 10.1016/j.jbc.2022.102400. Epub 2022 Aug 18. J Biol Chem. 2022. PMID: 35988645 Free PMC article.
-
Kinetic properties and substrate specificities of two recombinant human N-acetylglucosaminyltransferase-IV isozymes.Glycoconj J. 2006 Nov;23(7-8):473-80. doi: 10.1007/s10719-006-6216-3. Glycoconj J. 2006. PMID: 17006639
-
Regulation of human GnT-IV family activity by the lectin domain.Carbohydr Res. 2024 Nov;545:109285. doi: 10.1016/j.carres.2024.109285. Epub 2024 Oct 2. Carbohydr Res. 2024. PMID: 39369636 Review.
-
Targeted genetic inactivation of N-acetylglucosaminyltransferase-IVa impairs insulin secretion from pancreatic beta cells and evokes type 2 diabetes.Methods Enzymol. 2010;479:205-22. doi: 10.1016/S0076-6879(10)79012-1. Methods Enzymol. 2010. PMID: 20816168
-
Regulation of intracellular activity of N-glycan branching enzymes in mammals.J Biol Chem. 2024 Jul;300(7):107471. doi: 10.1016/j.jbc.2024.107471. Epub 2024 Jun 13. J Biol Chem. 2024. PMID: 38879010 Free PMC article. Review.
Cited by
-
Genetic glycoengineering in mammalian cells.J Biol Chem. 2021 Jan-Jun;296:100448. doi: 10.1016/j.jbc.2021.100448. Epub 2021 Feb 20. J Biol Chem. 2021. PMID: 33617880 Free PMC article. Review.
-
Density-dependent lectin-glycan interactions as a paradigm for conditional regulation by posttranslational modifications.Mol Cell Proteomics. 2013 Apr;12(4):913-20. doi: 10.1074/mcp.R112.026989. Epub 2013 Feb 1. Mol Cell Proteomics. 2013. PMID: 23378517 Free PMC article.
-
MGAT4A/Galectin9-Driven N-Glycosylation Aberration as a Promoting Mechanism for Poor Prognosis of Endometrial Cancer with TP53 Mutation.Adv Sci (Weinh). 2024 Dec;11(48):e2409764. doi: 10.1002/advs.202409764. Epub 2024 Nov 11. Adv Sci (Weinh). 2024. PMID: 39527463 Free PMC article.
-
N-Glycosylation.Adv Exp Med Biol. 2021;1325:3-24. doi: 10.1007/978-3-030-70115-4_1. Adv Exp Med Biol. 2021. PMID: 34495528
-
Suppression of cancer progression by MGAT1 shRNA knockdown.PLoS One. 2012;7(9):e43721. doi: 10.1371/journal.pone.0043721. Epub 2012 Sep 5. PLoS One. 2012. PMID: 22957033 Free PMC article.
References
-
- Asada M, Furukawa K, Segawa K, Endo T, Kobata A. Increased expression of highly branched N-glycans at cell surface is correlated with the malignant phenotypes of mouse tumor cells. Cancer Res. 1997;57:1073–1080. - PubMed
-
- Bhattacharyya R, Bhaumik M, Raju TS, Stanley P. Truncated, inactive N-acetylglucosaminyltransferase III (GlcNAc-TIII) induces neurological and other traits absent in mice that lack GlcNAc-TIII. J Biol Chem. 2002;277:26300–26309. - PubMed
-
- Ceroni A, Maass K, Geyer H, Geyer R, Dell A, Haslam SM. GlycoWorkbench: A tool for the computer-assisted annotation of mass spectra of glycans. J Proteome Res. 2008;7(4):1650–1659. - PubMed
-
- Comelli EM, Head SR, Gilmartin T, Whisenant T, Haslam SM, North SJ, Wong NW, Kudo T, Narimatsu H, Esko JD, et al. A focused microarray approach to functional glycomics: Transcriptional regulation of the glycome. Glycobiology. 2006;16(2):117–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
