

Intercellular adhesion molecule-4 and CD36 are implicated in the abnormal adhesiveness of sickle cell SAD mouse erythrocytes to endothelium
- PMID: 20015873
- PMCID: PMC2864378
- DOI: 10.3324/haematol.2009.017392
Intercellular adhesion molecule-4 and CD36 are implicated in the abnormal adhesiveness of sickle cell SAD mouse erythrocytes to endothelium
Abstract
Background: Abnormal adhesiveness of red blood cells to endothelium has been implicated in vaso-occlusive crisis of sickle cell disease. The present study examined whether the SAD mouse model exhibits the same abnormalities of red blood cell adhesion as those found in human sickle cell disease.
Design and methods: The repertoire of adhesive molecules on murine erythrocytes and bEnd.3 microvascular endothelial cells was determined by flow cytometry using monoclonal antibodies or by western blotting. Adhesion was investigated in dynamic conditions and measured at different shear stresses.
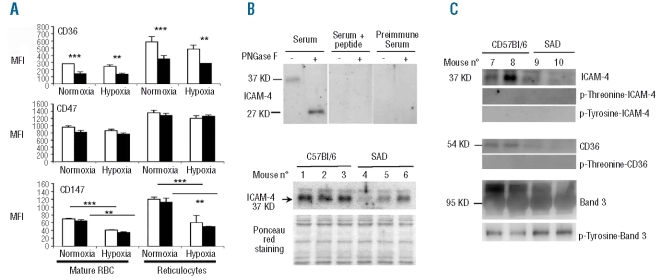
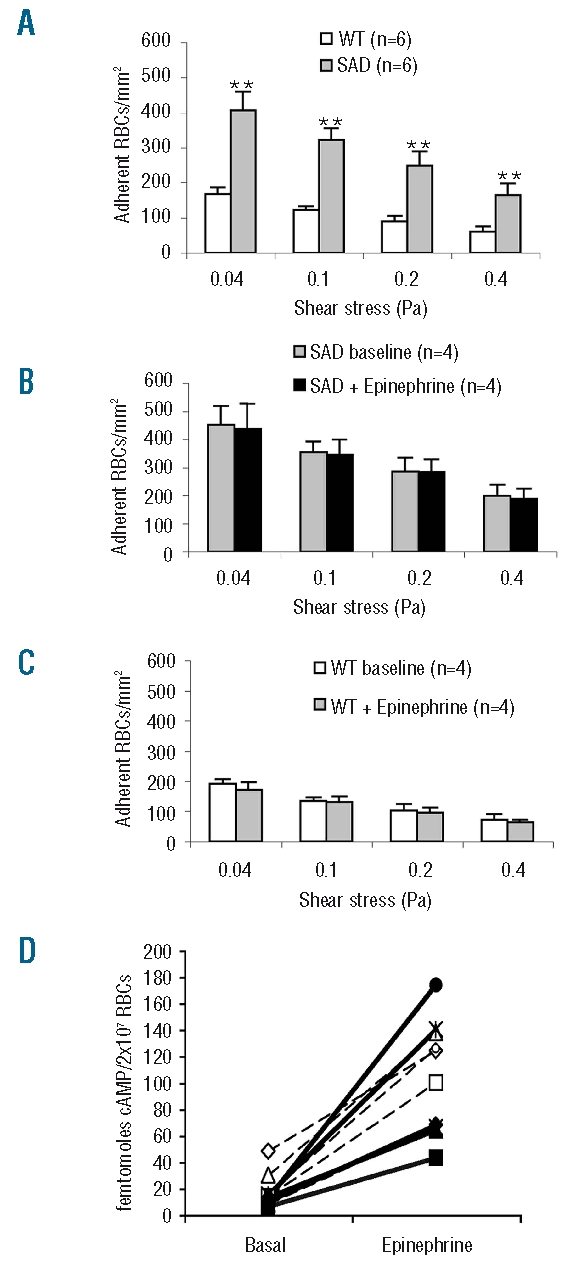
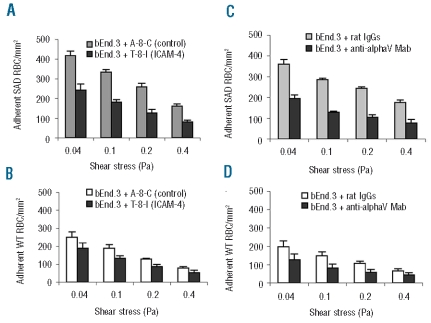
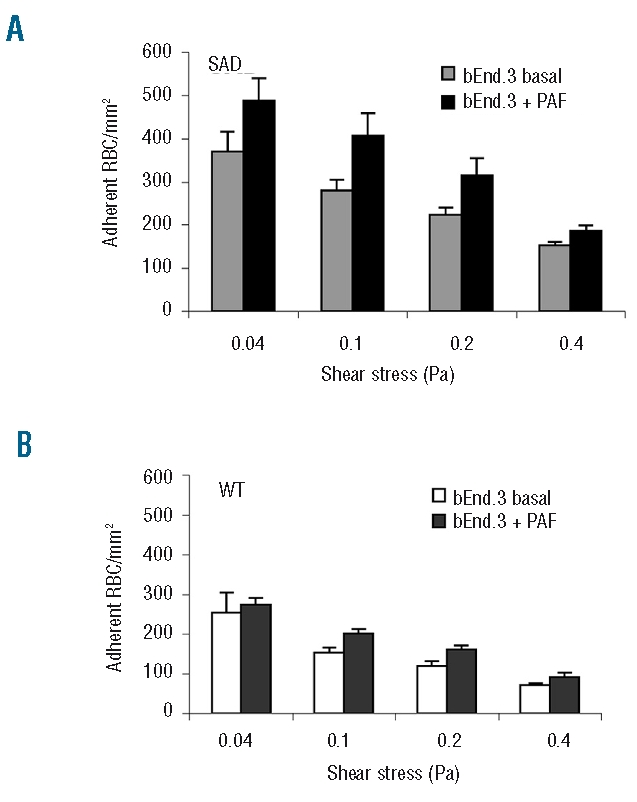
Results: CD36, CD47 and intercellular adhesion molecular-4, but not Lutheran blood group antigen/basal cell adhesion molecule, are present on mouse mature erythrocytes. alpha(4)beta(1) are not expressed on SAD and wild type reticulocytes. Endothelial bEnd.3 cells express alpha(V)beta(3), alpha(4)beta(1), CD47, vascular cell adhesion molecule-1, and Lutheran blood group antigen/basal cell adhesion molecule, but not CD36. Adhesion of SAD red cells is: (i) 2- to 3-fold higher than that of wild type red cells; (ii) further increased on platelet activating factor-activated endothelium; (iii) not stimulated by epinephrine; (iv) inhibited after treating the endothelium with a peptide reproducing one of the binding sequences of mouse intercellular adhesion molecular-4, or with mon-oclonal antibody against murine alpha(v) integrin; and (v) inhibited after pretreatment of red blood cells with anti-mouse CD36 monoclonal antibodies. The combination of treatments with intercellular adhesion molecular-4 peptide and anti-CD36 monoclonal antibodies eliminates excess adhesion of SAD red cells. The phosphorylation state of intercellular adhesion molecular-4 and CD36 is probably not involved in the over-adhesiveness of SAD erythrocytes.
Conclusions: Intercellular adhesion molecular-4/alpha(v)beta(3) and CD36/thrombospondin interactions might contribute to the abnormally high adhesiveness of SAD red cells. The SAD mouse is a valuable animal model for investigating adhesion processes of sickle cell disease.
Figures

Similar articles
-
Involvement of a CD47-dependent pathway in platelet adhesion on inflamed vascular endothelium under flow.Blood. 2003 Jun 15;101(12):4836-43. doi: 10.1182/blood-2002-11-3483. Epub 2003 Feb 27. Blood. 2003. PMID: 12609828
-
Phorbol ester stimulation increases sickle erythrocyte adherence to endothelium: a novel pathway involving alpha 4 beta 1 integrin receptors on sickle reticulocytes and fibronectin.Blood. 1996 Dec 1;88(11):4348-58. Blood. 1996. PMID: 8943872
-
Thrombospondin mediates adherence of CD36+ sickle reticulocytes to endothelial cells.Blood. 1992 Nov 15;80(10):2634-42. Blood. 1992. PMID: 1384794
-
Sickle red cell-endothelium interactions.Microcirculation. 2009 Jan;16(1):97-111. doi: 10.1080/10739680802279394. Microcirculation. 2009. PMID: 18720225 Free PMC article. Review.
-
[Erythrocyte adhesion to the vascular endothelium].Transfus Clin Biol. 1999 Dec;6(6):397-402. doi: 10.1016/s1246-7820(00)88984-3. Transfus Clin Biol. 1999. PMID: 10666797 Review. French.
Cited by
-
Red Blood Cells: Chasing Interactions.Front Physiol. 2019 Jul 31;10:945. doi: 10.3389/fphys.2019.00945. eCollection 2019. Front Physiol. 2019. PMID: 31417415 Free PMC article. Review.
-
Red Blood Cells: Tethering, Vesiculation, and Disease in Micro-Vascular Flow.Diagnostics (Basel). 2021 May 27;11(6):971. doi: 10.3390/diagnostics11060971. Diagnostics (Basel). 2021. PMID: 34072241 Free PMC article.
-
Differential modulation of adhesion molecule expression by hydroxycarbamide in human endothelial cells from the micro- and macrocirculation: potential implications in sickle cell disease vasoocclusive events.Haematologica. 2011 Apr;96(4):534-42. doi: 10.3324/haematol.2010.026740. Epub 2011 Jan 12. Haematologica. 2011. PMID: 21228039 Free PMC article.
-
Intercellular adhesion molecules (ICAMs) and spermatogenesis.Hum Reprod Update. 2013 Mar-Apr;19(2):167-86. doi: 10.1093/humupd/dms049. Epub 2013 Jan 3. Hum Reprod Update. 2013. PMID: 23287428 Free PMC article. Review.
-
Randomized phase 2 study of GMI-1070 in SCD: reduction in time to resolution of vaso-occlusive events and decreased opioid use.Blood. 2015 Apr 23;125(17):2656-64. doi: 10.1182/blood-2014-06-583351. Epub 2015 Mar 2. Blood. 2015. PMID: 25733584 Free PMC article. Clinical Trial.
References
-
- Hebbel RP, Boogaerts MA, Eaton JW, Steinberg MH. Erythrocyte adherence to endothelium in sickle-cell anemia. A possible determinant of disease severity. N Engl J Med. 1980;1;302(18):992–5. - PubMed
-
- Hebbel RP, Osarogiagbon R, Kaul D. The endothelial biology of sickle cell disease: inflammation and a chronic vasculopathy. Microcirculation. 2004;11(2):129–51. - PubMed
-
- Frenette PS. Sickle cell vasoocclusion: heterotypic, multicellular aggregations driven by leukocyte adhesion. Microcirculation. 2004;11(2):167–77. - PubMed
-
- Hines PC, Zen Q, Burney SN, Shea DA, Ataga KI, Orringer EP, et al. Novel epinephrine and cyclic AMP-mediated activation of BCAM/Lu-dependent sickle (SS) RBC adhesion. Blood. 2003;101(8):3281–7. - PubMed
-
- Zennadi R, Hines PC, De Castro LM, Cartron JP, Parise LV, Telen MJ. Epinephrine acts through erythroid signaling pathways to activate sickle cell adhesion to endothelium via LW-alphavbeta3 interactions. Blood. 2004;104(12):3774–81. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
