

Extracellular ubiquitin inhibits beta-AR-stimulated apoptosis in cardiac myocytes: role of GSK-3beta and mitochondrial pathways
- PMID: 20015977
- PMCID: PMC2836259
- DOI: 10.1093/cvr/cvp402
Extracellular ubiquitin inhibits beta-AR-stimulated apoptosis in cardiac myocytes: role of GSK-3beta and mitochondrial pathways
Abstract
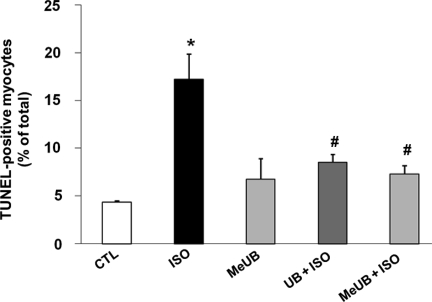
Aims: Beta-adrenergic receptor (beta-AR) stimulation induces apoptosis in adult rat ventricular myocytes (ARVMs) via the activation of glycogen synthase kinase-3beta (GSK-3beta) and mitochondrial pathways. However, beta-AR stimulation induces apoptosis only in a fraction ( approximately 15-20%) of ARVMs. We hypothesized that ARVMs may secrete/release a survival factor(s) which protects 80-85% of cells from apoptosis.
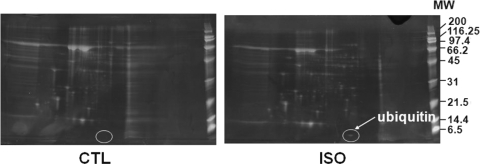
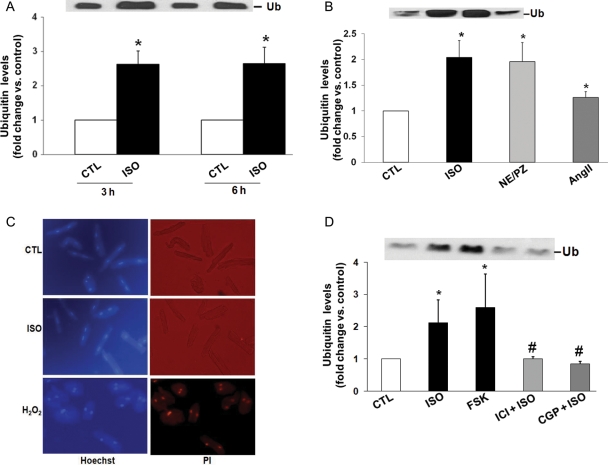
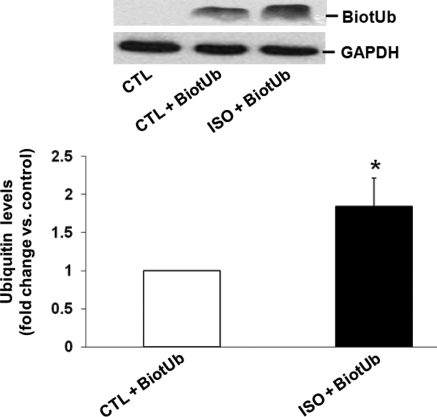
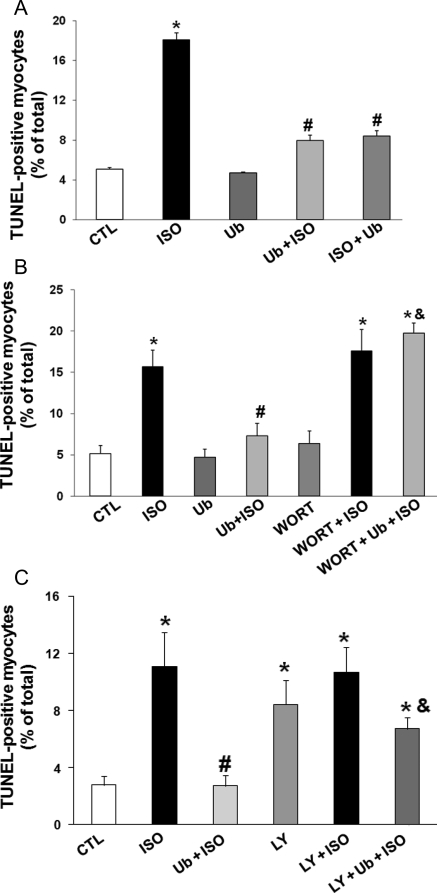
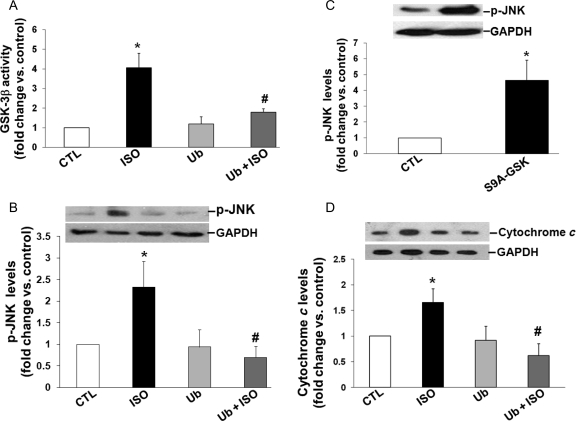
Methods and results: Using two-dimensional gel electrophoresis followed by MALDI TOF and MS/MS, we identified ubiquitin (Ub) in the conditioned media of ARVMs treated with beta-AR agonist (isoproterenol). Western blot analysis confirmed increased Ub levels in the conditioned media 3 and 6 h after beta-AR stimulation. Inhibition of beta1-AR and beta2-AR subtypes inhibited beta-AR-stimulated increases in extracellular levels of Ub, whereas activation of adenylyl cyclase using forskolin mimicked the effects of beta-AR stimulation. Incubation of cells with exogenous biotinylated Ub followed by western blot analysis of the cell lysates showed uptake of extracellular Ub into cells, which was found to be higher after beta-AR stimulation (1.9 +/- 0.4-fold; P < 0.05 vs. control, n = 6). Pre-treatment with Ub inhibited beta-AR-stimulated increases in apoptosis. Inhibition of phosphoinositide 3-kinase using wortmannin and LY-294002 prevented anti-apoptotic effects of extracellular Ub. Ub pre-treatment inhibited beta-AR-stimulated activation of GSK-3beta and c-Jun N-terminal kinase (JNK) and increases in the levels of cytosolic cytochrome c. The use of methylated Ub suggested that the anti-apoptotic effects of extracellular Ub are mediated via monoubiquitination.
Conclusion: beta-AR stimulation increases levels of Ub in the conditioned media. Extracellular Ub plays a protective role in beta-AR-stimulated apoptosis, possibly via the inactivation of GSK-3beta/JNK and mitochondrial pathways.
Figures
Comment in
-
Ubiquitin, a novel paracrine messenger of cardiac cell survival.Cardiovasc Res. 2010 Apr 1;86(1):1-3. doi: 10.1093/cvr/cvq026. Epub 2010 Jan 25. Cardiovasc Res. 2010. PMID: 20100704 Free PMC article. No abstract available.
References
-
- Andreka P, Nadhazi Z, Muzes G, Szantho G, Vandor L, Konya L, et al. Possible therapeutic targets in cardiac myocyte apoptosis. Curr Pharm Des. 2004;10:2445–2461. - PubMed
-
- Singh K, Xiao L, Remondino A, Sawyer DB, Colucci WS. Adrenergic regulation of cardiac myocyte apoptosis. J Cell Physiol. 2001;189:257–265. - PubMed
-
- Iwai-Kanai E, Hasegawa K, Araki M, Kakita T, Morimoto T, Sasayama S. Alpha- and beta-adrenergic pathways differentially regulate cell type-specific apoptosis in rat cardiac myocytes. Circulation. 1999;100:305–311. - PubMed
-
- Zaugg M, Xu W, Lucchinetti E, Shafiq SA, Jamali NZ, Siddiqui MA. Beta-adrenergic receptor subtypes differentially affect apoptosis in adult rat ventricular myocytes. Circulation. 2000;102:344–350. - PubMed
-
- Shizukuda Y, Buttrick PM, Geenen DL, Borczuk AC, Kitsis RN, Sonnenblick EH. Beta-adrenergic stimulation causes cardiocyte apoptosis: influence of tachycardia and hypertrophy. Am J Physiol. 1998;275:H961–H968. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
