

Targeted delivery of small interfering RNA to human dendritic cells to suppress dengue virus infection and associated proinflammatory cytokine production
- PMID: 20015996
- PMCID: PMC2820933
- DOI: 10.1128/JVI.02105-08
Targeted delivery of small interfering RNA to human dendritic cells to suppress dengue virus infection and associated proinflammatory cytokine production
Abstract
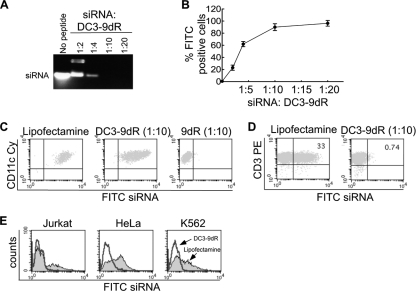
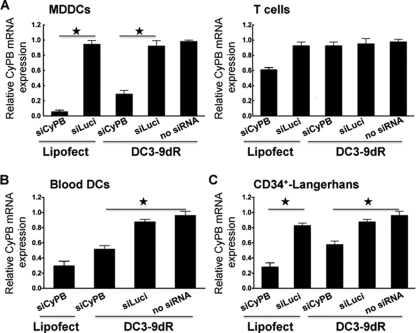
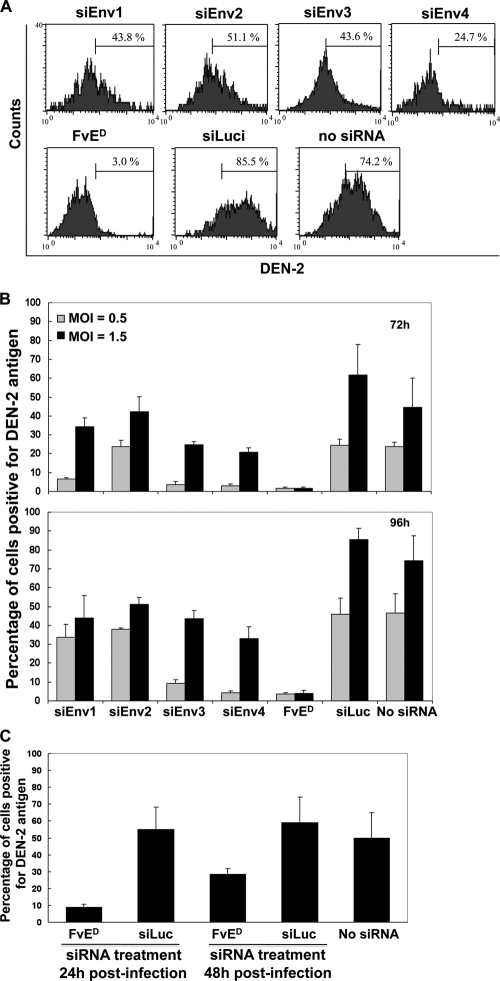
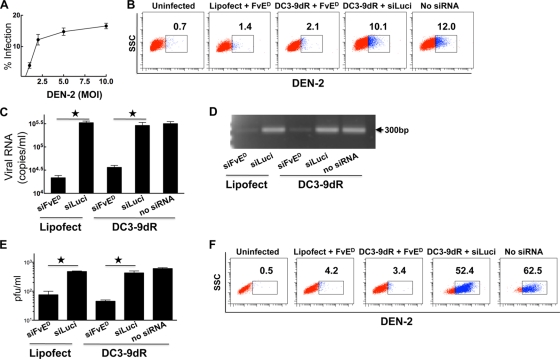
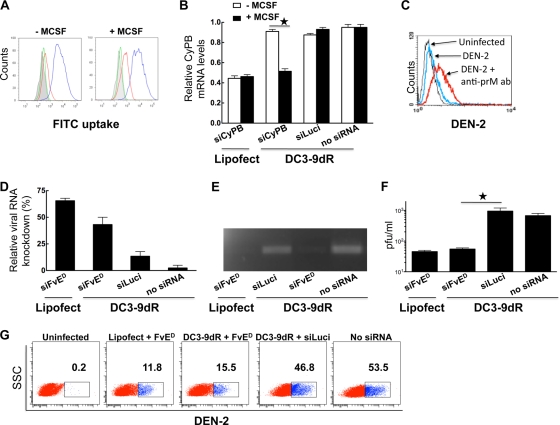
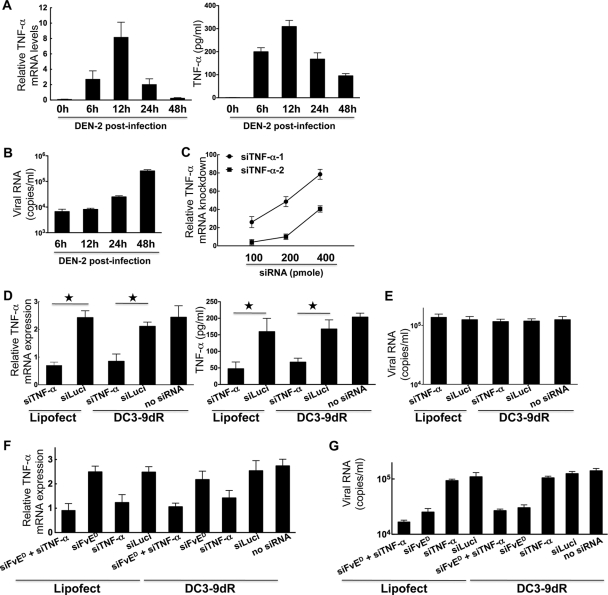
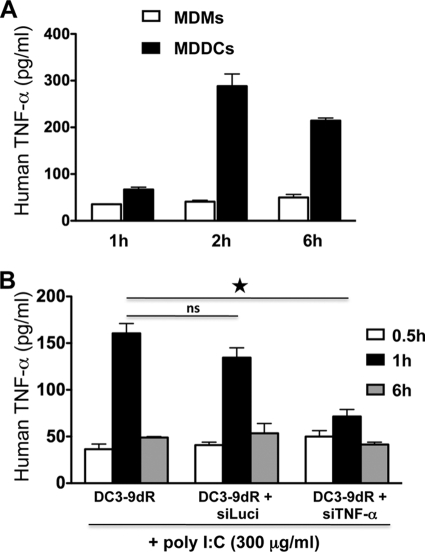
Dengue is a common arthropod-borne flaviviral infection in the tropics, for which there is no vaccine or specific antiviral drug. The infection is often associated with serious complications such as dengue hemorrhagic fever (DHF) or dengue shock syndrome (DSS), in which both viral and host factors have been implicated. RNA interference (RNAi) is a potent antiviral strategy and a potential therapeutic option for dengue if a feasible strategy can be developed for delivery of small interfering RNA (siRNA) to dendritic cells (DCs) and macrophages, the major in vivo targets of the virus and also the source of proinflammatory cytokines. Here we show that a dendritic cell-targeting 12-mer peptide (DC3) fused to nona-D-arginine (9dR) residues (DC3-9dR) delivers siRNA and knocks down endogenous gene expression in heterogenous DC subsets, (monocyte-derived DCs [MDDCs], CD34(+) hematopoietic stem cell [HSC])-derived Langerhans DCs, and peripheral blood DCs). Moreover, DC3-9dR-mediated delivery of siRNA targeting a highly conserved sequence in the dengue virus envelope gene (siFvE(D)) effectively suppressed dengue virus replication in MDDCs and macrophages. In addition, DC-specific delivery of siRNA targeting the acute-phase cytokine tumor necrosis factor alpha (TNF-alpha), which plays a major role in dengue pathogenesis, either alone or in combination with an antiviral siRNA, significantly reduced virus-induced production of the cytokine in MDDCs. Finally to validate the strategy in vivo, we tested the ability of the peptide to target human DCs in the NOD/SCID/IL-2Rgamma(-/-) mouse model engrafted with human CD34(+) hematopoietic stem cells (HuHSC mice). Treatment of mice by intravenous (i.v.) injection of DC3-9dR-complexed siRNA targeting TNF-alpha effectively suppressed poly(I:C)-induced TNF-alpha production by DCs. Thus, DC3-9dR can deliver siRNA to DCs both in vitro and in vivo, and this delivery approach holds promise as a therapeutic strategy to simultaneously suppress virus replication and curb virus-induced detrimental host immune responses in dengue infection.
Figures
Similar articles
-
Targeted delivery of siRNA to macrophages for anti-inflammatory treatment.Mol Ther. 2010 May;18(5):993-1001. doi: 10.1038/mt.2010.27. Epub 2010 Mar 9. Mol Ther. 2010. PMID: 20216529 Free PMC article.
-
Enhanced induction of HIV-specific cytotoxic T lymphocytes by dendritic cell-targeted delivery of SOCS-1 siRNA.Mol Ther. 2010 Nov;18(11):2028-37. doi: 10.1038/mt.2010.148. Epub 2010 Jul 20. Mol Ther. 2010. PMID: 20648001 Free PMC article.
-
Dengue-2 and yellow fever 17DD viruses infect human dendritic cells, resulting in an induction of activation markers, cytokines and chemokines and secretion of different TNF-α and IFN-α profiles.Mem Inst Oswaldo Cruz. 2011 Aug;106(5):594-605. doi: 10.1590/s0074-02762011000500012. Mem Inst Oswaldo Cruz. 2011. PMID: 21894381
-
The relationship of interacting immunological components in dengue pathogenesis.Virol J. 2009 Nov 27;6:211. doi: 10.1186/1743-422X-6-211. Virol J. 2009. PMID: 19941667 Free PMC article. Review.
-
Guiding dengue vaccine development using knowledge gained from the success of the yellow fever vaccine.Cell Mol Immunol. 2016 Jan;13(1):36-46. doi: 10.1038/cmi.2015.76. Epub 2015 Oct 5. Cell Mol Immunol. 2016. PMID: 26435066 Free PMC article. Review.
Cited by
-
Vitamin A or E and a catechin synergize as vaccine adjuvant to enhance immune responses in mice by induction of early interleukin-15 but not interleukin-1β responses.Immunology. 2016 Aug;148(4):352-62. doi: 10.1111/imm.12614. Epub 2016 Jun 22. Immunology. 2016. PMID: 27135790 Free PMC article.
-
RNA interference mediated inhibition of dengue virus multiplication and entry in HepG2 cells.PLoS One. 2012;7(3):e34060. doi: 10.1371/journal.pone.0034060. Epub 2012 Mar 23. PLoS One. 2012. PMID: 22457813 Free PMC article.
-
Arthropod-borne flaviviruses and RNA interference: seeking new approaches for antiviral therapy.Adv Virus Res. 2013;85:91-111. doi: 10.1016/B978-0-12-408116-1.00004-5. Adv Virus Res. 2013. PMID: 23439025 Free PMC article. Review.
-
Imino sugar glucosidase inhibitors as broadly active anti-filovirus agents.Emerg Microbes Infect. 2013 Nov;2(11):e77. doi: 10.1038/emi.2013.77. Epub 2013 Nov 20. Emerg Microbes Infect. 2013. PMID: 26038444 Free PMC article. Review.
-
Dengue Virus Non-Structural Protein 5 as a Versatile, Multi-Functional Effector in Host-Pathogen Interactions.Front Cell Infect Microbiol. 2021 Mar 18;11:574067. doi: 10.3389/fcimb.2021.574067. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 33816326 Free PMC article. Review.
References
-
- Bagasra, O. 2005. RNAi as an antiviral therapy. Expert Opin. Biol. Ther. 5:1463-1474. - PubMed
-
- Bai, F., T. Wang, U. Pal, F. Bao, L. H. Gould, and E. Fikrig. 2005. Use of RNA interference to prevent lethal murine West Nile virus infection. J. Infect. Dis. 191:1148-1154. - PubMed
-
- Basak, S. K., A. Harui, M. Stolina, S. Sharma, K. Mitani, S. M. Dubinett, and M. D. Roth. 2002. Increased dendritic cell number and function following continuous in vivo infusion of granulocyte macrophage-colony-stimulating factor and interleukin-4. Blood 99:2869-2879. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
