

Cardiac glycosides induce cell death in human cells by inhibiting general protein synthesis
- PMID: 20016840
- PMCID: PMC2788214
- DOI: 10.1371/journal.pone.0008292
Cardiac glycosides induce cell death in human cells by inhibiting general protein synthesis
Abstract
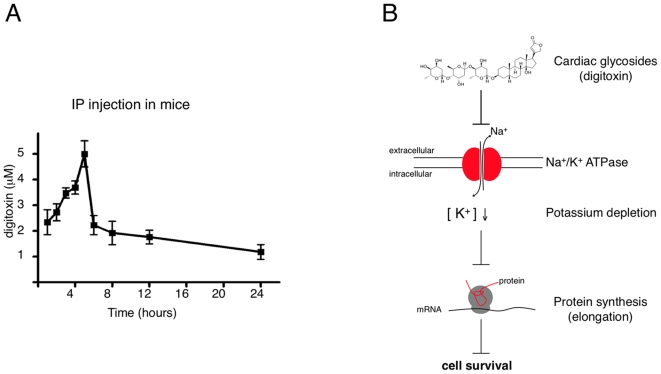
Background: Cardiac glycosides are Na(+)/K(+)-pump inhibitors widely used to treat heart failure. They are also highly cytotoxic, and studies have suggested specific anti-tumor activity leading to current clinical trials in cancer patients. However, a definitive demonstration of this putative anti-cancer activity and the underlying molecular mechanism has remained elusive.
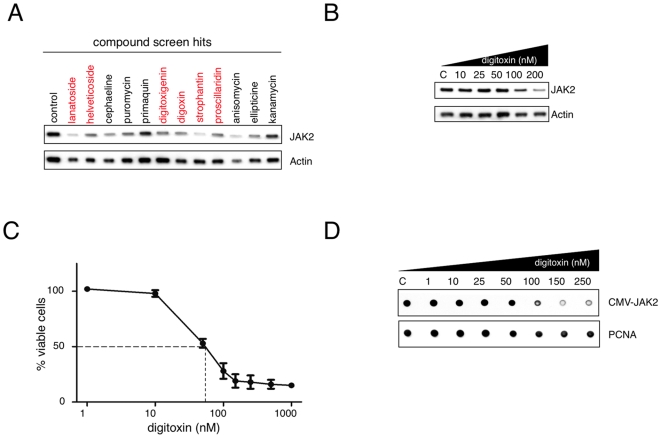
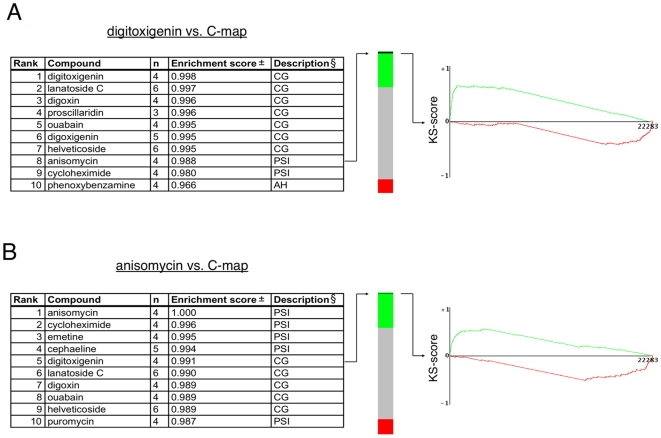
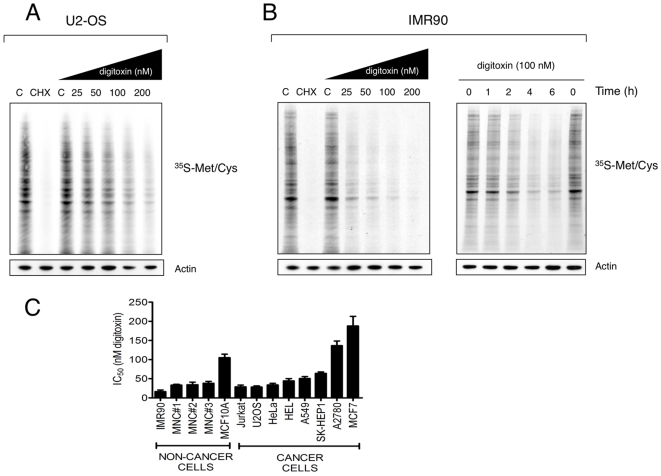
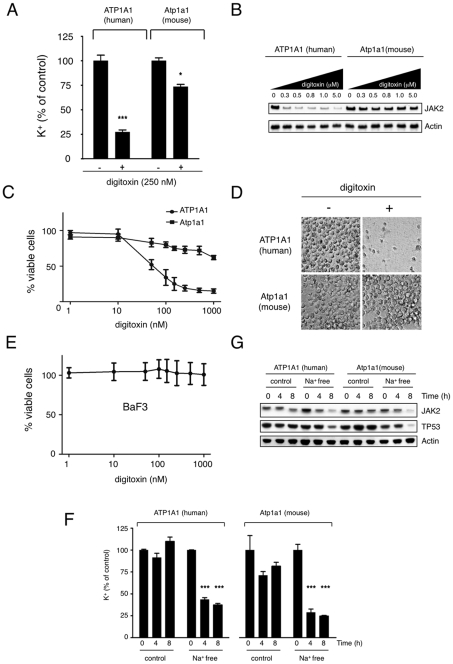
Methodology/principal findings: Using an unbiased transcriptomics approach, we found that cardiac glycosides inhibit general protein synthesis. Protein synthesis inhibition and cytotoxicity were not specific for cancer cells as they were observed in both primary and cancer cell lines. These effects were dependent on the Na(+)/K(+)-pump as they were rescued by expression of a cardiac glycoside-resistant Na(+)/K(+)-pump. Unlike human cells, rodent cells are largely resistant to cardiac glycosides in vitro and mice were found to tolerate extremely high levels.
Conclusions/significance: The physiological difference between human and mouse explains the previously observed sensitivity of human cancer cells in mouse xenograft experiments. Thus, published mouse xenograft models used to support anti-tumor activity for these drugs require reevaluation. Our finding that cardiac glycosides inhibit protein synthesis provides a mechanism for the cytotoxicity of CGs and raises concerns about ongoing clinical trials to test CGs as anti-cancer agents in humans.
Conflict of interest statement
Figures
References
-
- Therien AG, Blostein R. Mechanisms of sodium pump regulation. Am J Physiol Cell Physiol. 2000;279:C541–566. - PubMed
-
- Prassas I, Diamandis EP. Novel therapeutic applications of cardiac glycosides. Nat Rev Drug Discov. 2008;7:926–935. - PubMed
-
- Mijatovic T, Van Quaquebeke E, Delest B, Debeir O, Darro F, et al. Cardiotonic steroids on the road to anti-cancer therapy. Biochim Biophys Acta. 2007;1776:32–57. - PubMed
-
- Zavareh RB, Lau KS, Hurren R, Datti A, Ashline DJ, et al. Inhibition of the sodium/potassium ATPase impairs N-glycan expression and function. Cancer Res. 2008;68:6688–6697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
