

Stimulation of nonselective amino acid export by glutamine dumper proteins
- PMID: 20018597
- PMCID: PMC2815850
- DOI: 10.1104/pp.109.151746
Stimulation of nonselective amino acid export by glutamine dumper proteins
Abstract
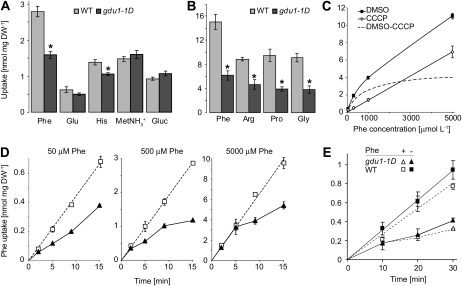
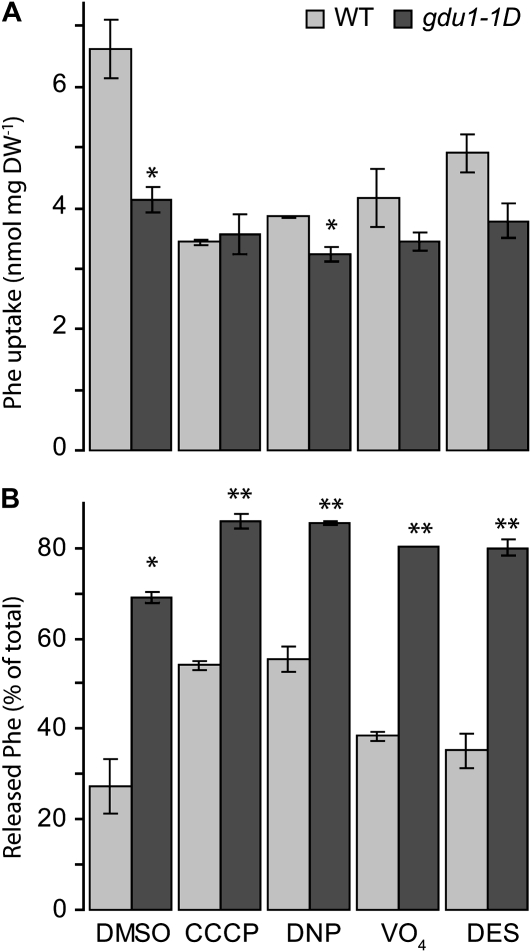
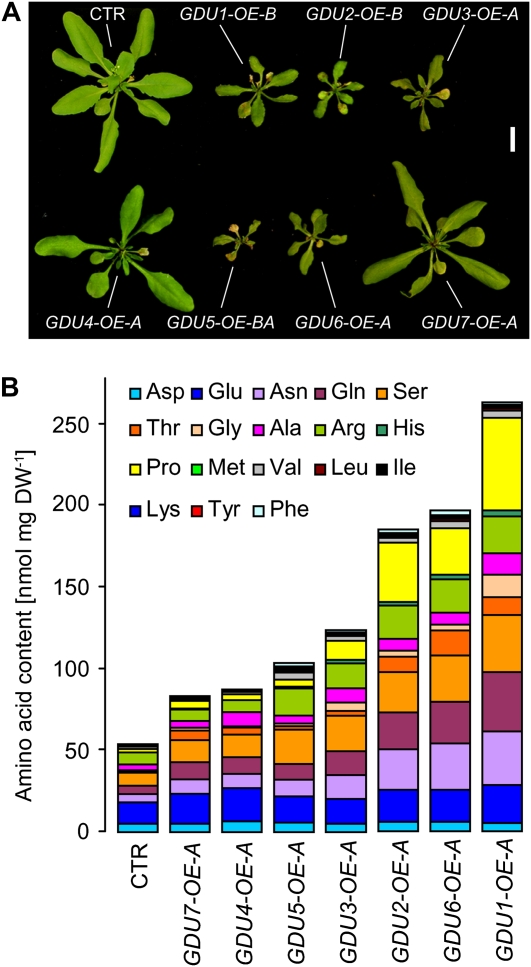
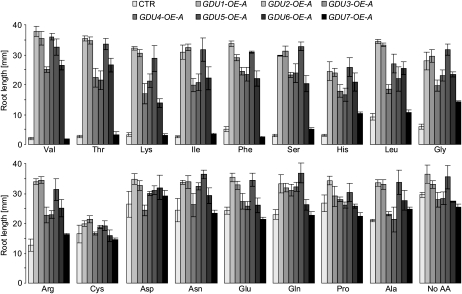
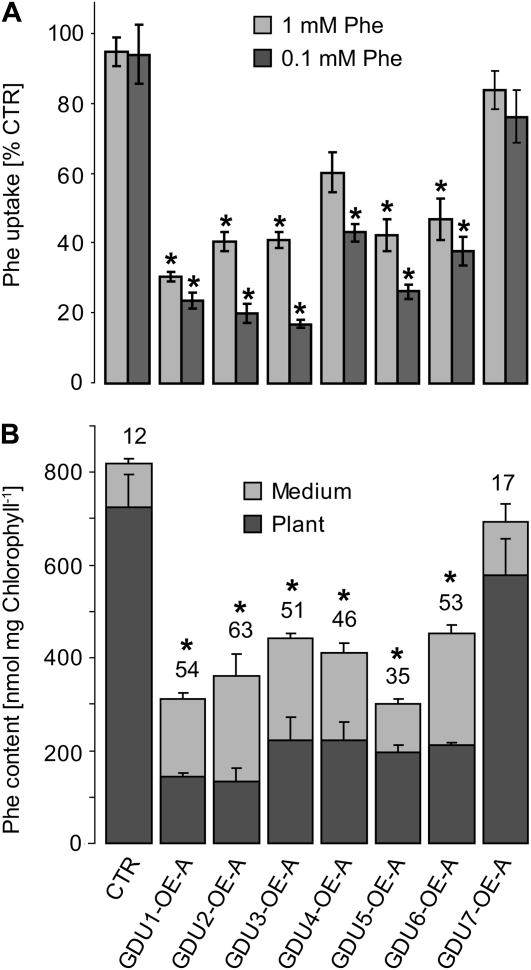
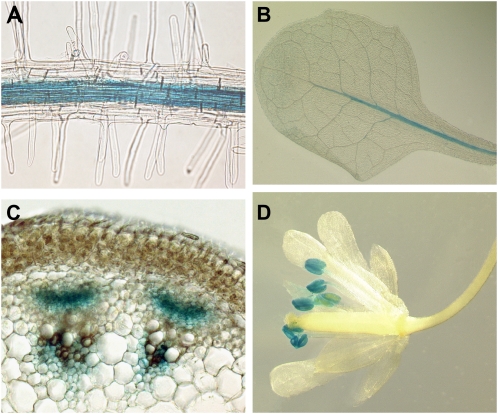
Phloem and xylem transport of amino acids involves two steps: export from one cell type to the apoplasm, and subsequent import into adjacent cells. High-affinity import is mediated by proton/amino acid cotransporters, while the mechanism of export remains unclear. Enhanced expression of the plant-specific type I membrane protein Glutamine Dumper1 (GDU1) has previously been shown to induce the secretion of glutamine from hydathodes and increased amino acid content in leaf apoplasm and xylem sap. In this work, tolerance to low concentrations of amino acids and transport analyses using radiolabeled amino acids demonstrate that net amino acid uptake is reduced in the glutamine-secreting GDU1 overexpressor gdu1-1D. The net uptake rate of phenylalanine decreased over time, and amino acid net efflux was increased in gdu1-1D compared with the wild type, indicating increased amino acid export from cells. Independence of the export from proton gradients and ATP suggests that overexpression of GDU1 affects a passive export system. Each of the seven Arabidopsis (Arabidopsis thaliana) GDU genes led to similar phenotypes, including increased efflux of a wide spectrum of amino acids. Differences in expression profiles and functional properties suggested that the GDU genes fulfill different roles in roots, vasculature, and reproductive organs. Taken together, the GDUs appear to stimulate amino acid export by activating nonselective amino acid facilitators.
Figures

Similar articles
-
Overexpression of GLUTAMINE DUMPER1 leads to hypersecretion of glutamine from Hydathodes of Arabidopsis leaves.Plant Cell. 2004 Jul;16(7):1827-40. doi: 10.1105/tpc.021642. Epub 2004 Jun 18. Plant Cell. 2004. PMID: 15208395 Free PMC article.
-
The ubiquitin E3 ligase LOSS OF GDU2 is required for GLUTAMINE DUMPER1-induced amino acid secretion in Arabidopsis.Plant Physiol. 2012 Apr;158(4):1628-42. doi: 10.1104/pp.111.191965. Epub 2012 Jan 30. Plant Physiol. 2012. PMID: 22291198 Free PMC article.
-
Control of Amino Acid Homeostasis by a Ubiquitin Ligase-Coactivator Protein Complex.J Biol Chem. 2017 Mar 3;292(9):3827-3840. doi: 10.1074/jbc.M116.766469. Epub 2017 Jan 18. J Biol Chem. 2017. PMID: 28100770 Free PMC article.
-
Suppressor mutations in the Glutamine Dumper1 protein dissociate disturbance in amino acid transport from other characteristics of the Gdu1D phenotype.Front Plant Sci. 2015 Aug 4;6:593. doi: 10.3389/fpls.2015.00593. eCollection 2015. Front Plant Sci. 2015. PMID: 26300894 Free PMC article.
-
Amino acid export in plants: a missing link in nitrogen cycling.Mol Plant. 2011 May;4(3):453-63. doi: 10.1093/mp/ssr003. Epub 2011 Feb 15. Mol Plant. 2011. PMID: 21324969 Free PMC article. Review.
Cited by
-
A spatiotemporal transcriptomic network dynamically modulates stalk development in maize.Plant Biotechnol J. 2022 Dec;20(12):2313-2331. doi: 10.1111/pbi.13909. Epub 2022 Sep 7. Plant Biotechnol J. 2022. PMID: 36070002 Free PMC article.
-
Siliques are Red1 from Arabidopsis acts as a bidirectional amino acid transporter that is crucial for the amino acid homeostasis of siliques.Plant Physiol. 2012 Apr;158(4):1643-55. doi: 10.1104/pp.111.192583. Epub 2012 Feb 6. Plant Physiol. 2012. PMID: 22312005 Free PMC article.
-
Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli.Front Plant Sci. 2019 Feb 21;10:157. doi: 10.3389/fpls.2019.00157. eCollection 2019. Front Plant Sci. 2019. PMID: 30881364 Free PMC article. Review.
-
Defective cytokinin signaling reprograms lipid and flavonoid gene-to-metabolite networks to mitigate high salinity in Arabidopsis.Proc Natl Acad Sci U S A. 2021 Nov 30;118(48):e2105021118. doi: 10.1073/pnas.2105021118. Proc Natl Acad Sci U S A. 2021. PMID: 34815339 Free PMC article.
-
Intrusive Growth of Phloem Fibers in Flax Stem: Integrated Analysis of miRNA and mRNA Expression Profiles.Plants (Basel). 2019 Feb 19;8(2):47. doi: 10.3390/plants8020047. Plants (Basel). 2019. PMID: 30791461 Free PMC article.
References
-
- Bonner CA, Williams DS, Aldrich HC, Jensen RA (1996) Antagonism by L-glutamine of toxicity and growth inhibition caused by other amino acids in suspension cultures of Nicotiana sylvestris. Plant Sci 113 43–58
-
- Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16 735–743 - PubMed
-
- Dundar E (2009) Multiple GUS expression patterns of a single Arabidopsis gene. Ann Appl Biol 154 33–41
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
