

MEF9, an E-subclass pentatricopeptide repeat protein, is required for an RNA editing event in the nad7 transcript in mitochondria of Arabidopsis
- PMID: 20018598
- PMCID: PMC2815870
- DOI: 10.1104/pp.109.151175
MEF9, an E-subclass pentatricopeptide repeat protein, is required for an RNA editing event in the nad7 transcript in mitochondria of Arabidopsis
Abstract
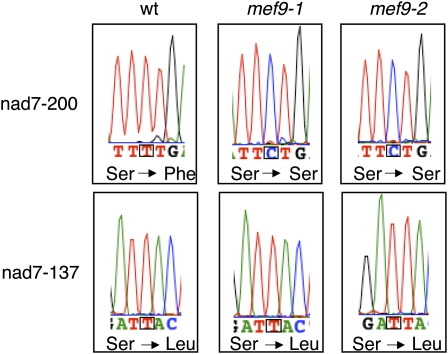
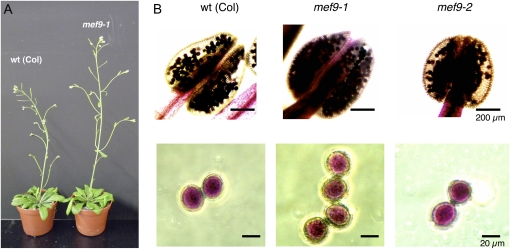
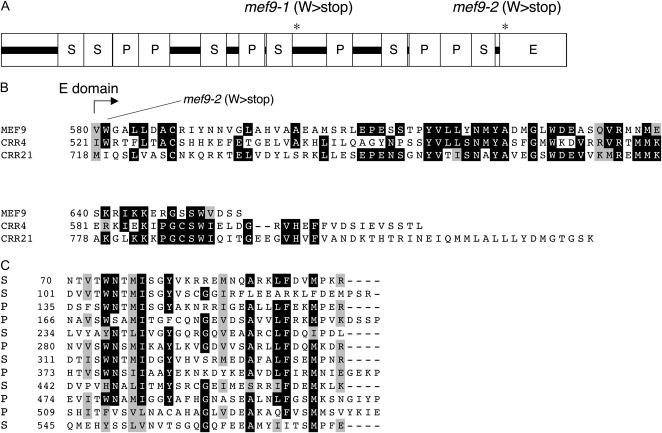
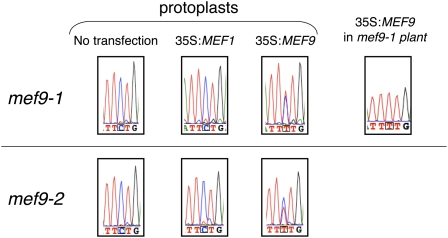
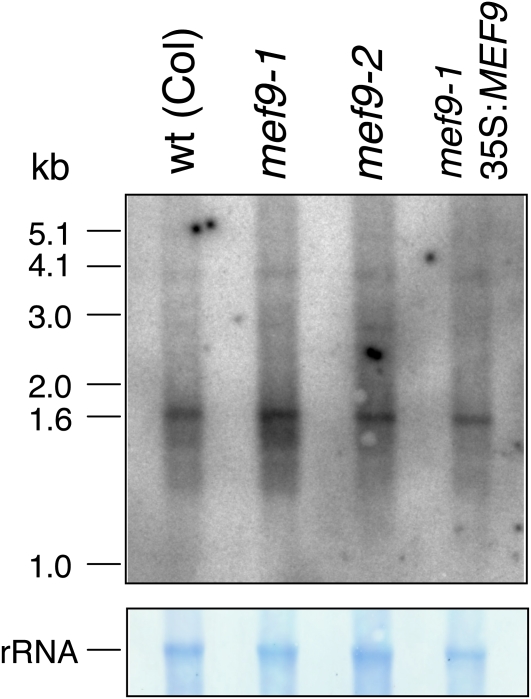
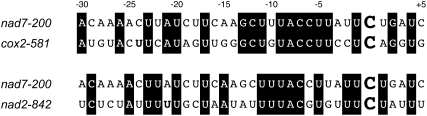
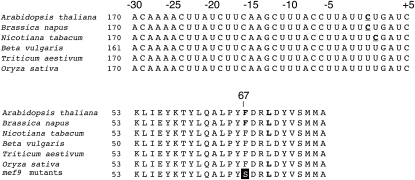
RNA editing in plants alters specific nucleotides from C to U in mRNAs in plastids and in mitochondria. I here characterize the nuclear gene MITOCHONDRIAL EDITING FACTOR9 (MEF9) that is required for RNA editing of the site nad7-200 in the nad7 mitochondrial mRNA in Arabidopsis (Arabidopsis thaliana). The MEF9 protein belongs to the E subfamily of pentatricopeptide repeat proteins and unlike the three previously identified mitochondrial editing factors MEF1 and MEF11 in Arabidopsis and OGR1 in rice (Oryza sativa) does not contain a DYW C-terminal domain. In addition, the E domain is incomplete, but seems to be functionally required, since one of the two independent EMS mutants encodes a MEF9 protein truncated by a stop codon at the beginning of the E domain. In both mutant plants premature stop codons in MEF9 inactivate RNA editing at site nad7-200. The homozygous mutant plants are viable and develop rather normally. The lack of RNA editing at site nad7-200 thus seems to be tolerated although this editing event is conserved in most plant species or the genomic sequence already codes for a T at this position, resulting in a generally conserved amino acid codon.
Figures
References
-
- Alexander MP (1969) Differential staining of aborted and nonaborted pollen. Stain Technol 44 117–122 - PubMed
-
- Andrés C, Lurin C, Small ID (2007) The multifarious roles of PPR proteins in plant mitochondrial gene expression. Physiol Plant 129 14–22
-
- Chateigner-Boutin AL, Ramos-Vega M, Guevara-García A, Andrés C, de la Luz Gutiérrez-Nava M, Cantero A, Delannoy E, Jiménez LF, Lurin C, Small ID, et al (2008) CLB19, a pentatricopeptide repeat protein required for editing of rpoA and clpP chloroplast transcripts. Plant J 56 590–602 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
