

Terminal deoxynucleotidyl transferase is required for an optimal response to the polysaccharide α-1,3 dextran
- PMID: 20018621
- PMCID: PMC4710363
- DOI: 10.4049/jimmunol.0902791
Terminal deoxynucleotidyl transferase is required for an optimal response to the polysaccharide α-1,3 dextran
Abstract
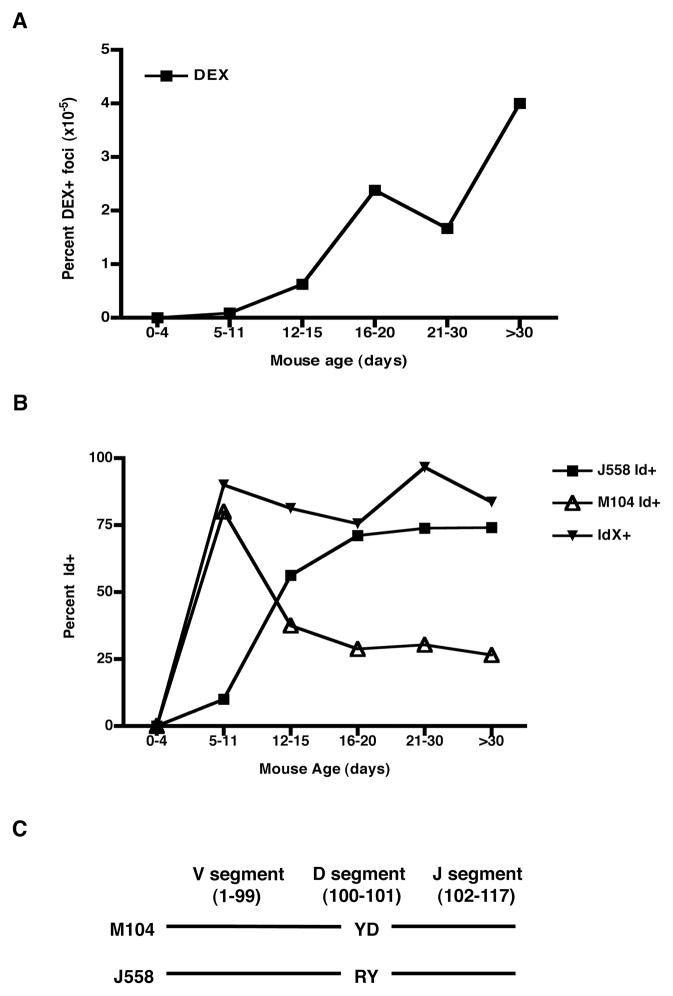
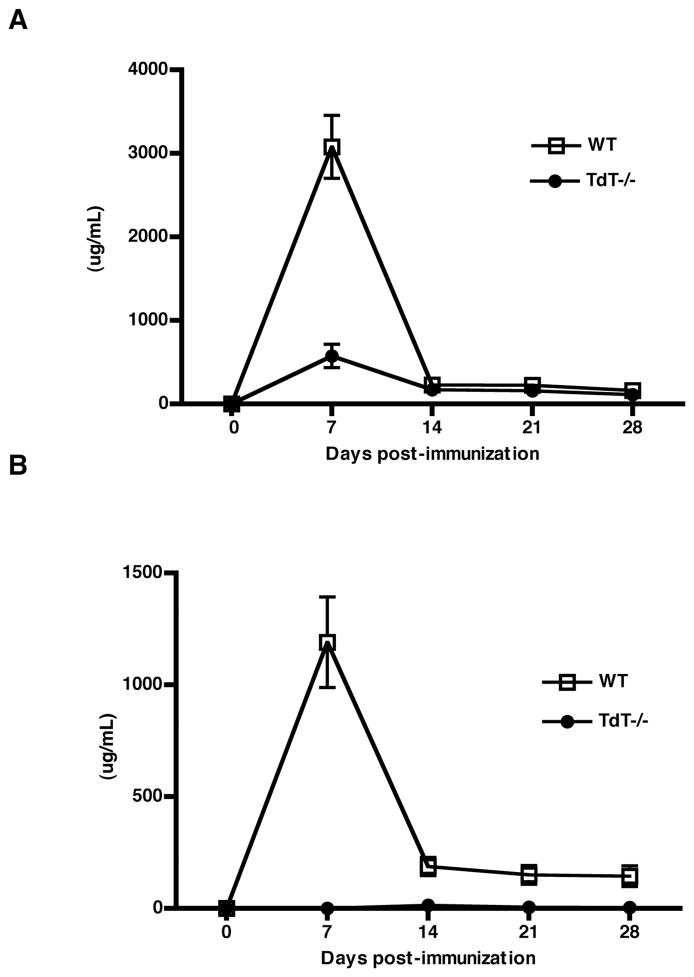
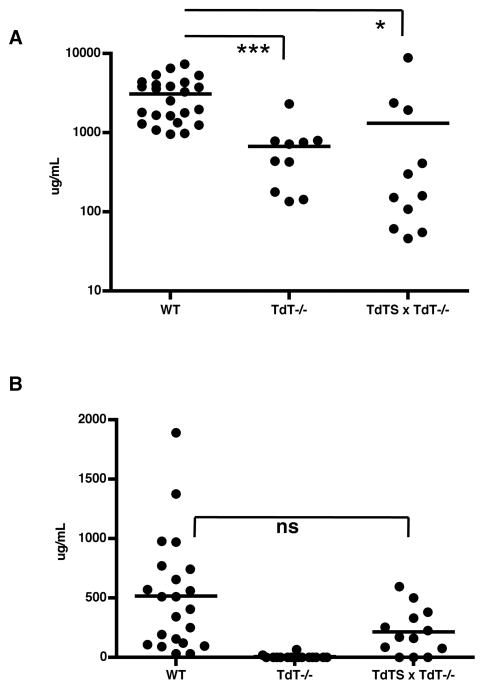
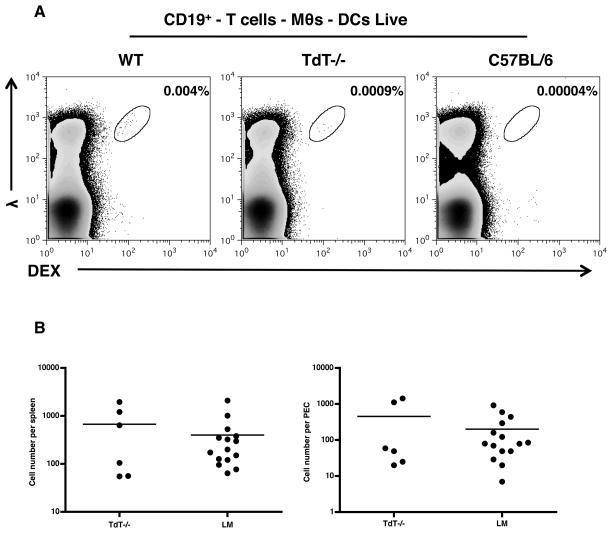
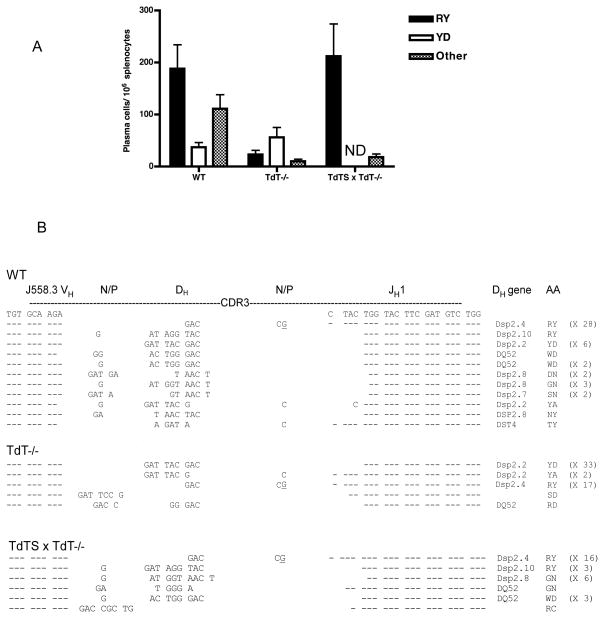
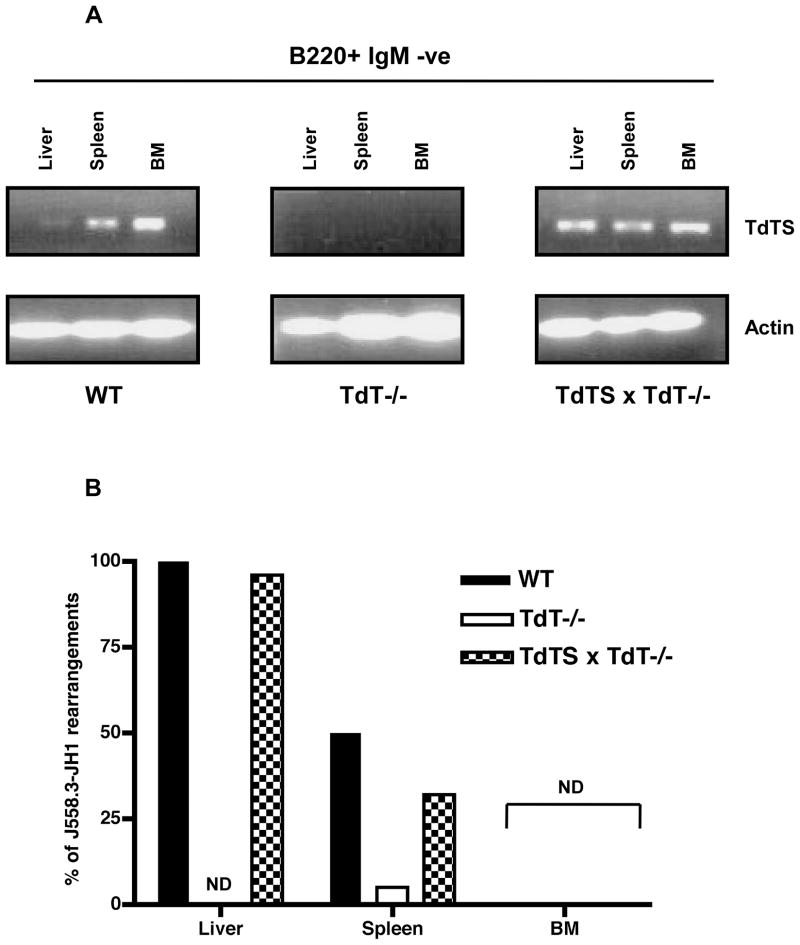
An understanding of Ab responses to polysaccharides associated with pathogenic microorganisms is of importance for improving vaccine design, especially in neonates that respond poorly to these types of Ags. In this study, we have investigated the role of the lymphoid-specific enzyme TdT in generating B cell clones responsive to alpha-1,3 dextran (DEX). TdT is a DNA polymerase that plays a major role in generating diversity of lymphocyte AgRs during V(D)J recombination. In this study, we show that the DEX-specific Ab response is lower, and the dominant DEX-specific J558 idiotype (Id) is not detected in TdT(-/-) mice when compared with wild-type (WT) BALB/c mice. Nucleotide sequencing of H chain CDR3s of DEX-specific plasmablasts, sorted postimmunization, showed that TdT(-/-) mice generate a lower frequency of the predominant adult molecularly determined clone J558. Complementation of TdT expression in TdT(-/-) mice by early forced expression of the short splice variant of TdT-restored WT proportions of J558 Id+ clones and also abrogated the development of the minor M104E Id+ clones. J558 Id V(D)J rearrangements are detected as early as 7 d after birth in IgM-negative B cell precursors in the liver and spleen of WT and TdT-transgenic mice but not in TdT(-/-) mice. These data show that TdT is essential for the generation of the predominant higher-affinity DEX-responsive J558 clone.
Figures
Similar articles
-
Absence of N addition facilitates B cell development, but impairs immune responses.Immunogenetics. 2011 Sep;63(9):599-609. doi: 10.1007/s00251-011-0543-7. Epub 2011 Jun 10. Immunogenetics. 2011. PMID: 21660592 Free PMC article.
-
Limiting CDR-H3 diversity abrogates the antibody response to the bacterial polysaccharide α 1→3 dextran.J Immunol. 2011 Jul 15;187(2):879-86. doi: 10.4049/jimmunol.1100957. Epub 2011 Jun 15. J Immunol. 2011. PMID: 21677133 Free PMC article.
-
The M603 idiotype is lost in the response to phosphocholine in terminal deoxynucleotidyl transferase-deficient mice.Eur J Immunol. 2002 Apr;32(4):1139-46. doi: 10.1002/1521-4141(200204)32:4<1139::AID-IMMU1139>3.0.CO;2-E. Eur J Immunol. 2002. PMID: 11932921
-
Terminal deoxynucleotidyl transferase and repertoire development.Immunol Rev. 2000 Jun;175:150-7. Immunol Rev. 2000. PMID: 10933600 Review.
-
Mice lacking terminal deoxynucleotidyl transferase: adult mice with a fetal antigen receptor repertoire.Immunol Rev. 1995 Dec;148:201-19. doi: 10.1111/j.1600-065x.1995.tb00099.x. Immunol Rev. 1995. PMID: 8825288 Review.
Cited by
-
Differential idiotype utilization for the in vivo type 14 capsular polysaccharide-specific Ig responses to intact Streptococcus pneumoniae versus a pneumococcal conjugate vaccine.J Immunol. 2012 Jul 15;189(2):575-86. doi: 10.4049/jimmunol.1200599. Epub 2012 Jun 15. J Immunol. 2012. PMID: 22706079 Free PMC article.
-
Absence of N addition facilitates B cell development, but impairs immune responses.Immunogenetics. 2011 Sep;63(9):599-609. doi: 10.1007/s00251-011-0543-7. Epub 2011 Jun 10. Immunogenetics. 2011. PMID: 21660592 Free PMC article.
-
Limiting CDR-H3 diversity abrogates the antibody response to the bacterial polysaccharide α 1→3 dextran.J Immunol. 2011 Jul 15;187(2):879-86. doi: 10.4049/jimmunol.1100957. Epub 2011 Jun 15. J Immunol. 2011. PMID: 21677133 Free PMC article.
-
Pulmonary α-1,3-Glucan-Specific IgA-Secreting B Cells Suppress the Development of Cockroach Allergy.J Immunol. 2016 Oct 15;197(8):3175-3187. doi: 10.4049/jimmunol.1601039. Epub 2016 Aug 31. J Immunol. 2016. PMID: 27581173 Free PMC article.
-
Terminal Deoxynucleotidyl Transferase Is Not Required for Antibody Response to Polysaccharide Vaccines against Streptococcus pneumoniae and Salmonella enterica Serovar Typhi.Infect Immun. 2018 Aug 22;86(9):e00211-18. doi: 10.1128/IAI.00211-18. Print 2018 Sep. Infect Immun. 2018. PMID: 29967094 Free PMC article.
References
-
- Comstock LE, Kasper DL. Bacterial glycans: key mediators of diverse host immune responses. Cell. 2006;126:847–850. - PubMed
-
- Mazmanian SK, Kasper DL. The love-hate relationship between bacterial polysaccharides and the host immune system. Nat Rev Immunol. 2006;6:849–858. - PubMed
-
- Lesinski GB, Westerink MA. Vaccines against polysaccharide antigens. Curr Drug Targets Infect Disord. 2001;1:325–334. - PubMed
-
- Weintraub A. Immunology of bacterial polysaccharide antigens. Carbohydr Res. 2003;338:2539–2547. - PubMed
-
- Vos Q, Lees A, Wu ZQ, Snapper CM, Mond JJ. B-cell activation by T-cell-independent type 2 antigens as an integral part of the humoral immune response to pathogenic microorganisms. Immunol Rev. 2000;176:154–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
