

A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity
- PMID: 20018686
- PMCID: PMC2806711
- DOI: 10.1073/pnas.0909705107
A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity
Abstract
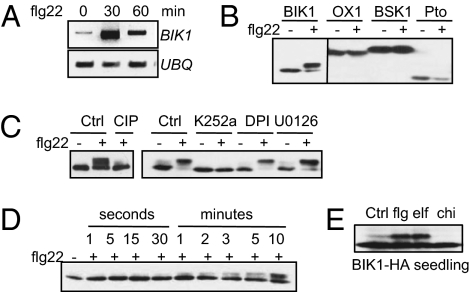
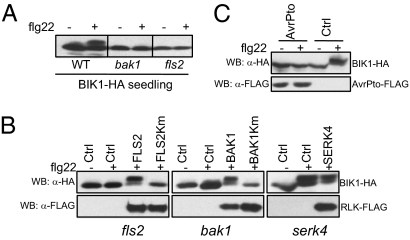
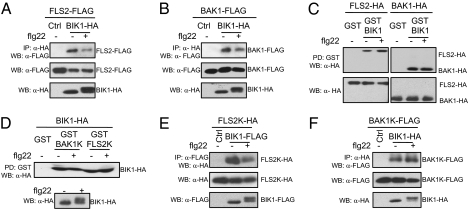
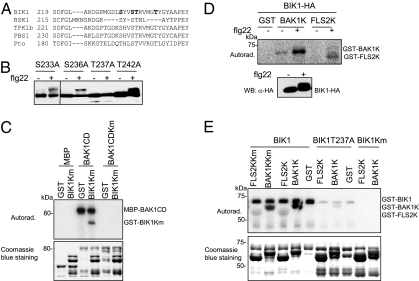
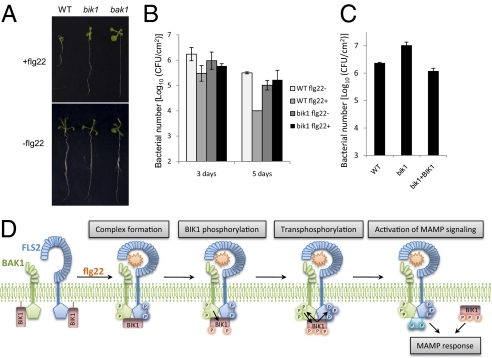
Plants and animals rely on innate immunity to prevent infections by detection of microbe-associated molecular patterns (MAMPs) through pattern-recognition receptors (PRRs). The plant PRR FLS2, a leucine-rich repeat-receptor kinase, recognizes bacterial flagellin and initiates immune signaling by association with another leucine-rich repeat-receptor-like kinase, BAK1. It remains unknown how the FLS2/BAK1 receptor complex activates intracellular signaling cascades. Here we identified the receptor-like cytoplasmic kinase BIK1 that is rapidly phosphorylated upon flagellin perception, depending on both FLS2 and BAK1. BIK1 associates with FLS2 and BAK1 in vivo and in vitro. BIK1 is phosphorylated by BAK1, and BIK1 also directly phosphorylates BAK1 and FLS2 in vitro. The flagellin phosphorylation site Thr(237) of BIK1 is required for its phosphorylation on BAK1 and FLS2, suggesting that BIK1 is likely first phosphorylated upon flagellin perception and subsequently transphosphorylates FLS2/BAK1 to propagate flagellin signaling. Importantly, bik1 mutants are compromised in diverse flagellin-mediated responses and immunity to the nonpathogenic bacterial infection. Thus, BIK1 is an essential component in MAMP signal transduction, which links the MAMP receptor complex to downstream intracellular signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
-
- Chisholm ST, Coaker G, Day B, Staskawicz BJ. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell. 2006;124:803–814. - PubMed
-
- Boller T, Felix G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol. 2009;60:379–406. - PubMed
-
- Bent AF, Mackey D. Elicitors, effectors, and R genes: The new paradigm and a lifetime supply of questions. Annu Rev Phytopathol. 2007;45:399–436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
