

Involvement of the spliceosomal U4 small nuclear RNA in heterochromatic gene silencing at fission yeast centromeres
- PMID: 20018856
- PMCID: PMC2820790
- DOI: 10.1074/jbc.M109.074393
Involvement of the spliceosomal U4 small nuclear RNA in heterochromatic gene silencing at fission yeast centromeres
Abstract
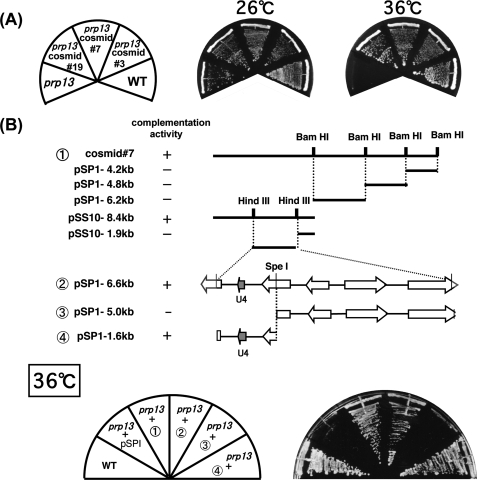
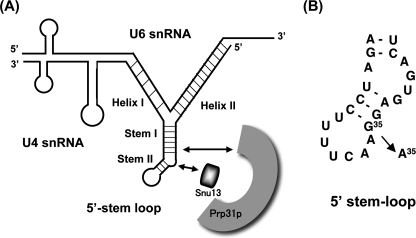
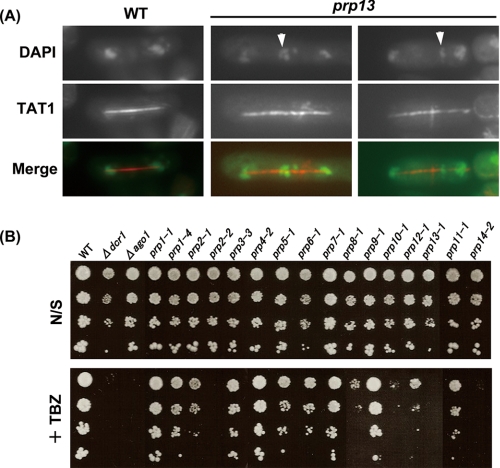
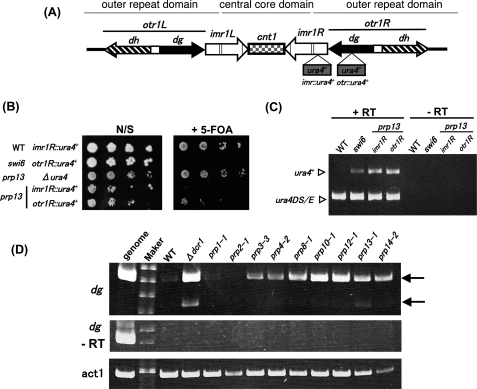
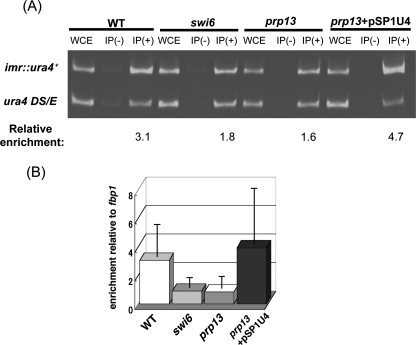
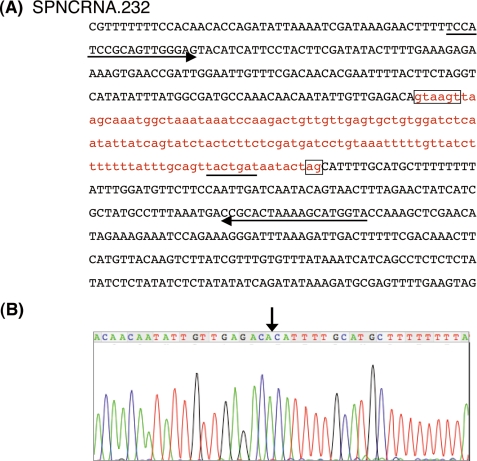
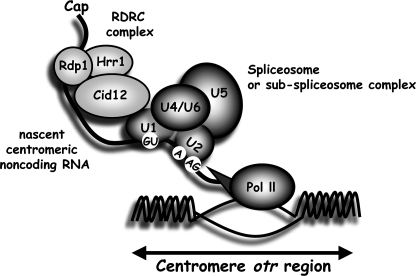
prp13-1 is one of the mutants isolated in a screen for defective pre-mRNA splicing at a nonpermissive temperature in fission yeast Schizosaccharomyces pombe. We cloned the prp13(+) gene and found that it encodes U4 small nuclear RNA (snRNA) involved in the assembly of the spliceosome. The prp13-1 mutant produced elongated cells, a phenotype similar to cell division cycle mutants, and displays a high incidence of lagging chromosomes on anaphase spindles. The mutant is hypersensitive to the microtubule-destabilizing drug thiabendazole, supporting that prp13-1 has a defect in chromosomal segregation. We found that the prp13-1 mutation resulted in expression of the ura4(+) gene inserted in the pericentromeric heterochromatin region and reduced recruitment of the heterochromatin protein Swi6p to that region, indicating defects in the formation of pericentromeric heterochromatin, which is essential for the segregation of chromosomes, in prp13-1. The formation of centromeric heterochromatin is induced by the RNA interference (RNAi) system in S. pombe. In prp13-1, the processing of centromeric noncoding RNAs to siRNAs, which direct the heterochromatin formation, was impaired and unprocessed noncoding RNAs were accumulated. These results suggest that U4 snRNA is required for the RNAi-directed heterochromatic gene silencing at the centromeres. In relation to the linkage between the spliceosomal U4 snRNA and the RNAi-directed formation of heterochromatin, we identified a mRNA-type intron in the centromeric noncoding RNAs. We propose a model in which the assembly of the spliceosome or a sub-spliceosome complex on the intron-containing centromeric noncoding RNAs facilitates the RNAi-directed formation of heterochromatin at centromeres, through interaction with the RNA-directed RNA polymerase complex.
Figures

Similar articles
-
The chromatin-remodeling factor FACT contributes to centromeric heterochromatin independently of RNAi.Curr Biol. 2007 Jul 17;17(14):1219-24. doi: 10.1016/j.cub.2007.06.028. Epub 2007 Jul 5. Curr Biol. 2007. PMID: 17614284 Free PMC article.
-
Studies on the mechanism of RNAi-dependent heterochromatin assembly.Cold Spring Harb Symp Quant Biol. 2006;71:461-71. doi: 10.1101/sqb.2006.71.044. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381328 Review.
-
Interaction of APC/C-E3 ligase with Swi6/HP1 and Clr4/Suv39 in heterochromatin assembly in fission yeast.J Biol Chem. 2009 Mar 13;284(11):7165-76. doi: 10.1074/jbc.M806461200. Epub 2008 Dec 30. J Biol Chem. 2009. PMID: 19117951 Free PMC article.
-
Heterochromatin and RNAi are required to establish CENP-A chromatin at centromeres.Science. 2008 Jan 4;319(5859):94-7. doi: 10.1126/science.1150944. Science. 2008. PMID: 18174443 Free PMC article.
-
On the connection between RNAi and heterochromatin at centromeres.Cold Spring Harb Symp Quant Biol. 2010;75:275-83. doi: 10.1101/sqb.2010.75.024. Epub 2011 Feb 2. Cold Spring Harb Symp Quant Biol. 2010. PMID: 21289046 Review.
Cited by
-
Splicing regulation by long noncoding RNAs.Nucleic Acids Res. 2018 Mar 16;46(5):2169-2184. doi: 10.1093/nar/gky095. Nucleic Acids Res. 2018. PMID: 29425321 Free PMC article. Review.
-
Red5 and three nuclear pore components are essential for efficient suppression of specific mRNAs during vegetative growth of fission yeast.Nucleic Acids Res. 2013 Jul;41(13):6674-86. doi: 10.1093/nar/gkt363. Epub 2013 May 8. Nucleic Acids Res. 2013. PMID: 23658229 Free PMC article.
-
RNA and epigenetic silencing: insight from fission yeast.Dev Growth Differ. 2012 Jan;54(1):129-41. doi: 10.1111/j.1440-169X.2011.01310.x. Epub 2011 Dec 12. Dev Growth Differ. 2012. PMID: 22150237 Free PMC article. Review.
-
RNA Splicing Factors and RNA-Directed DNA Methylation.Biology (Basel). 2014 Mar 26;3(2):243-54. doi: 10.3390/biology3020243. Biology (Basel). 2014. PMID: 24833507 Free PMC article.
-
The PRP6-like splicing factor STA1 is involved in RNA-directed DNA methylation by facilitating the production of Pol V-dependent scaffold RNAs.Nucleic Acids Res. 2013 Oct;41(18):8489-502. doi: 10.1093/nar/gkt639. Epub 2013 Jul 22. Nucleic Acids Res. 2013. PMID: 23877244 Free PMC article.
References
-
- Jurica M. S., Moore M. J. (2003) Mol. Cell 12, 5–14 - PubMed
-
- Burge C. B., Tuschl T., Sharp P. A. (1999) in The RNA Word (Gesteland R. F., Cech T. R., Atkins J. F. eds) pp. 525–560, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York
-
- Valadkhan S. (2005) Curr. Opin. Chem. Biol. 9, 603–608 - PubMed
-
- Staley J. P., Guthrie C. (1998) Cell 92, 315–326 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
