

Mitochondrial dynamics regulate the RIG-I-like receptor antiviral pathway
- PMID: 20019757
- PMCID: PMC2828750
- DOI: 10.1038/embor.2009.258
Mitochondrial dynamics regulate the RIG-I-like receptor antiviral pathway
Abstract
The intracellular retinoic acid-inducible gene I-like receptors (RLRs) sense viral ribonucleic acid and signal through the mitochondrial protein mitochondrial antiviral signalling (MAVS) to trigger the production of type I interferons and proinflammatory cytokines. In this study, we report that RLR activation promotes elongation of the mitochondrial network. Mimicking this elongation enhances signalling downstream from MAVS and favours the binding of MAVS to stimulator of interferon genes, an endoplasmic reticulum (ER) protein involved in the RLR pathway. By contrast, enforced mitochondrial fragmentation dampens signalling and reduces the association between both proteins. Our finding that MAVS is associated with a pool of mitofusin 1, a protein of the mitochondrial fusion machinery, suggests that MAVS is capable of regulating mitochondrial dynamics to facilitate the mitochondria-ER association required for signal transduction. Importantly, we observed that viral mitochondria-localized inhibitor of apoptosis, a cytomegalovirus (CMV) antiapoptotic protein that promotes mitochondrial fragmentation, inhibits signalling downstream from MAVS, suggesting a possible new immune modulation strategy of the CMV.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
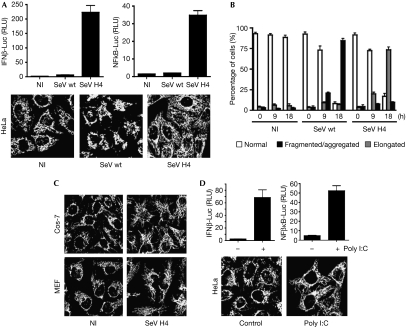
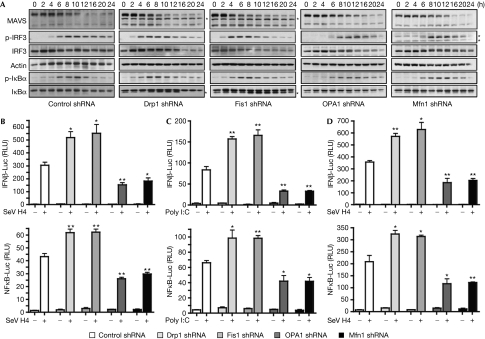
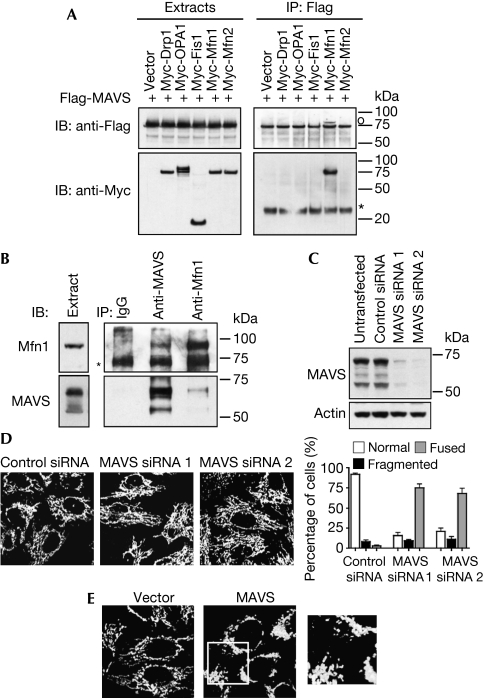
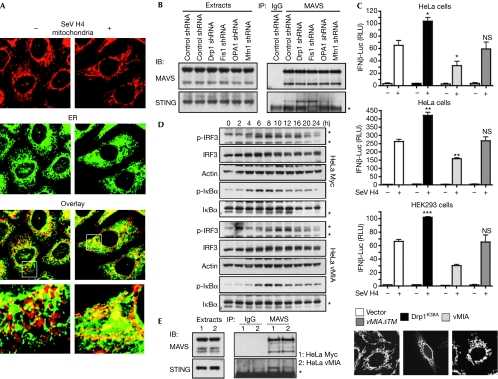
References
-
- Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124: 783–801 - PubMed
-
- Arnoult D, Carneiro L, Tattoli I, Girardin SE (2009) The role of mitochondria in cellular defense against microbial infection. Semin Immunol 21: 223–232 - PubMed
-
- Chan DC (2006) Mitochondria: dynamic organelles in disease, aging, and development. Cell 125: 1241–1252 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
