

Nano-motion dynamics are determined by surface-tethered selectin mechanokinetics and bond formation
- PMID: 20019797
- PMCID: PMC2787012
- DOI: 10.1371/journal.pcbi.1000612
Nano-motion dynamics are determined by surface-tethered selectin mechanokinetics and bond formation
Abstract
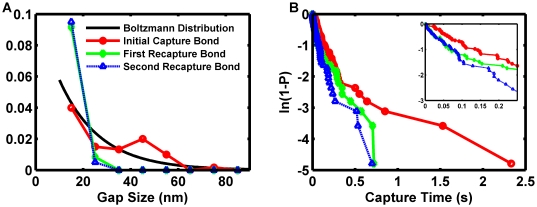
The interaction of proteins at cellular interfaces is critical for many biological processes, from intercellular signaling to cell adhesion. For example, the selectin family of adhesion receptors plays a critical role in trafficking during inflammation and immunosurveillance. Quantitative measurements of binding rates between surface-constrained proteins elicit insight into how molecular structural details and post-translational modifications contribute to function. However, nano-scale transport effects can obfuscate measurements in experimental assays. We constructed a biophysical simulation of the motion of a rigid microsphere coated with biomolecular adhesion receptors in shearing flow undergoing thermal motion. The simulation enabled in silico investigation of the effects of kinetic force dependence, molecular deformation, grouping adhesion receptors into clusters, surface-constrained bond formation, and nano-scale vertical transport on outputs that directly map to observable motions. Simulations recreated the jerky, discrete stop-and-go motions observed in P-selectin/PSGL-1 microbead assays with physiologic ligand densities. Motion statistics tied detailed simulated motion data to experimentally reported quantities. New deductions about biomolecular function for P-selectin/PSGL-1 interactions were made. Distributing adhesive forces among P-selectin/PSGL-1 molecules closely grouped in clusters was necessary to achieve bond lifetimes observed in microbead assays. Initial, capturing bond formation effectively occurred across the entire molecular contour length. However, subsequent rebinding events were enhanced by the reduced separation distance following the initial capture. The result demonstrates that vertical transport can contribute to an enhancement in the apparent bond formation rate. A detailed analysis of in silico motions prompted the proposition of wobble autocorrelation as an indicator of two-dimensional function. Insight into two-dimensional bond formation gained from flow cell assays might therefore be important to understand processes involving extended cellular interactions, such as immunological synapse formation. A biologically informative in silico system was created with minimal, high-confidence inputs. Incorporating random effects in surface separation through thermal motion enabled new deductions of the effects of surface-constrained biomolecular function. Important molecular information is embedded in the patterns and statistics of motion.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Nano-to-micro scale dynamics of P-selectin detachment from leukocyte interfaces. III. Numerical simulation of tethering under flow.Biophys J. 2005 Mar;88(3):1676-83. doi: 10.1529/biophysj.104.051805. Epub 2004 Dec 1. Biophys J. 2005. PMID: 15574709 Free PMC article.
-
The state diagram for cell adhesion mediated by two receptors.Biophys J. 2003 Apr;84(4):2671-90. doi: 10.1016/S0006-3495(03)75073-5. Biophys J. 2003. PMID: 12668476 Free PMC article.
-
Simulation and Analysis of Tethering Behavior of Neutrophils with Pseudopods.PLoS One. 2015 Jun 19;10(6):e0128378. doi: 10.1371/journal.pone.0128378. eCollection 2015. PLoS One. 2015. PMID: 26091091 Free PMC article.
-
Selectins promote tumor metastasis.Semin Cancer Biol. 2010 Jun;20(3):169-77. doi: 10.1016/j.semcancer.2010.04.005. Epub 2010 May 7. Semin Cancer Biol. 2010. PMID: 20452433 Review.
-
PSGL-1 function in immunity and steady state homeostasis.Immunol Rev. 2009 Jul;230(1):75-96. doi: 10.1111/j.1600-065X.2009.00797.x. Immunol Rev. 2009. PMID: 19594630 Review.
Cited by
-
Minimal encounter time and separation determine ligand-receptor binding in cell adhesion.Biophys J. 2011 Jun 8;100(11):2642-51. doi: 10.1016/j.bpj.2011.04.011. Biophys J. 2011. PMID: 21641309 Free PMC article.
-
Biomechanics of leukocyte rolling.Biorheology. 2011;48(1):1-35. doi: 10.3233/BIR-2011-0579. Biorheology. 2011. PMID: 21515934 Free PMC article. Review.
-
Single-molecule imaging and microfluidic platform reveal molecular mechanisms of leukemic cell rolling.Commun Biol. 2021 Jul 14;4(1):868. doi: 10.1038/s42003-021-02398-2. Commun Biol. 2021. PMID: 34262131 Free PMC article.
-
Multiscale computational models of complex biological systems.Annu Rev Biomed Eng. 2013;15:137-54. doi: 10.1146/annurev-bioeng-071811-150104. Epub 2013 Apr 29. Annu Rev Biomed Eng. 2013. PMID: 23642247 Free PMC article. Review.
-
Leukocyte rolling on engineered nanodot surfaces.Micro Nano Lett. 2011 May;6(5):301-305. doi: 10.1049/mnl.2011.0184. Epub 2011 May 30. Micro Nano Lett. 2011. PMID: 39512362 Free PMC article.
References
-
- Bell GI. Models for the specific adhesion of cells to cells. Science. 1978;200:618–627. - PubMed
-
- Lawrence MB. Selectin-carbohydrate interactions in shear flow. Curr Opin Chem Biol. 1999;3:659–664. - PubMed
-
- Thomas WE, Trintchina E, Forero M, Vogel V, Sokurenko EV. Bacterial adhesion to target cells enhanced by shear force. Cell. 2002;109:913–923. - PubMed
-
- Marshall BT, Long M, Piper JW, Yago T, McEver RP, et al. Direct observation of catch bonds involving cell-adhesion molecules. Nature. 2003;423:190–193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
