

The Noc-domain containing C-terminus of Noc4p mediates both formation of the Noc4p-Nop14p submodule and its incorporation into the SSU processome
- PMID: 20019888
- PMCID: PMC2794458
- DOI: 10.1371/journal.pone.0008370
The Noc-domain containing C-terminus of Noc4p mediates both formation of the Noc4p-Nop14p submodule and its incorporation into the SSU processome
Abstract
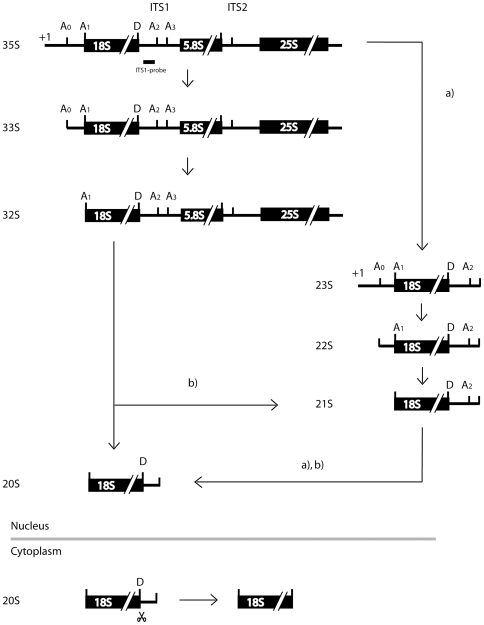
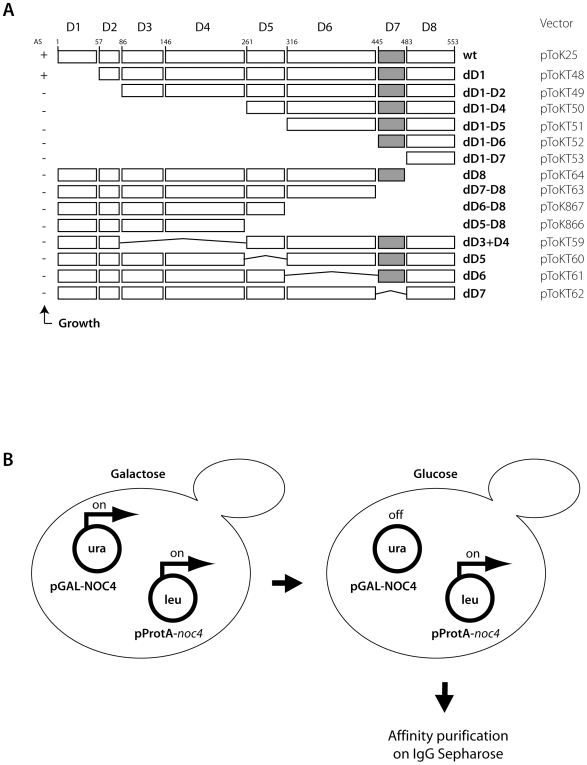
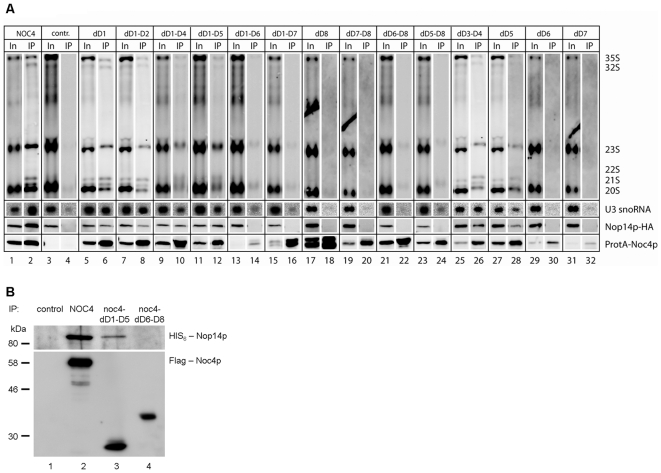
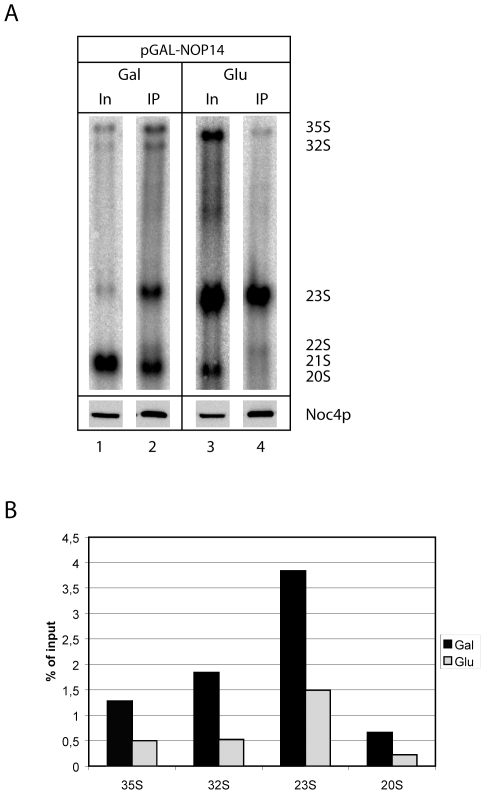
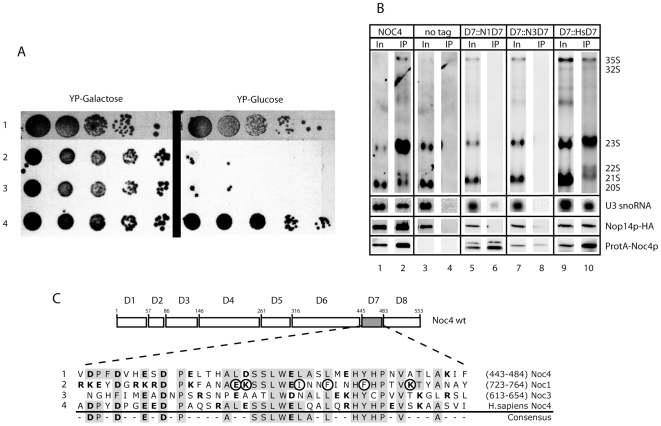
Noc1p, Noc3p and Noc4p are eukaryotic proteins which play essential roles in yeast ribosome biogenesis and contain a homologous stretch of about 45 aminoacids (Noc-domain) of unknown function. Yeast Noc4p is a component of the small ribosomal subunit (SSU) processome, can be isolated as a stable Noc4p-Nop14p SSU-processome submodule from yeast cells, and is required for nuclear steps of small ribosomal subunit rRNA maturation. We expressed a series of mutated alleles of NOC4 in yeast cells and analysed whether the corresponding protein variants support vegetative growth, interact with Nop14p, and are incorporated into the SSU-processome. The data reveal that the essential C-terminus of Noc4p which contains 237 aminoacids including the Noc-domain represents a protein-protein interaction module. It is required and sufficient for its association with Nop14p and several nuclear precursors of the small ribosomal subunit. The N-terminal Noc4-part seems to be targeted to pre-ribosomes via the C-terminus of Noc4p and plays there an essential role in SSU-processome function. Replacement of the Noc4p-Noc-domain by its homologues Noc1p-counterpart results in a hybrid Noc4p variant which fails to associate with Nop14p and pre-ribosomes. On the other hand, exchange of 6 amino acids in the Noc1-Noc-domain of this hybrid Noc4p protein is sufficient to restore its essential in vivo functions. These data suggest that Noc-domains of Noc1p and Noc4p share a common structural backbone in which diverging amino acids play crucial roles in mediating specific regulated interactions. Our analysis allows us to distinguish between different functions of certain domains within Noc4p and contribute to the understanding of how incorporation of Noc4p into ribosomal precursors is coupled to rRNA processing and maturation of the small ribosomal subunit.
Conflict of interest statement
Figures
Similar articles
-
Interrelationships between yeast ribosomal protein assembly events and transient ribosome biogenesis factors interactions in early pre-ribosomes.PLoS One. 2012;7(3):e32552. doi: 10.1371/journal.pone.0032552. Epub 2012 Mar 14. PLoS One. 2012. PMID: 22431976 Free PMC article.
-
A Noc complex specifically involved in the formation and nuclear export of ribosomal 40 S subunits.J Biol Chem. 2003 Feb 7;278(6):4072-81. doi: 10.1074/jbc.M208898200. Epub 2002 Nov 20. J Biol Chem. 2003. PMID: 12446671
-
Components of an interdependent unit within the SSU processome regulate and mediate its activity.Mol Cell Biol. 2002 Oct;22(20):7258-67. doi: 10.1128/MCB.22.20.7258-7267.2002. Mol Cell Biol. 2002. PMID: 12242301 Free PMC article.
-
Assembly and structure of the SSU processome-a nucleolar precursor of the small ribosomal subunit.Curr Opin Struct Biol. 2018 Apr;49:85-93. doi: 10.1016/j.sbi.2018.01.008. Epub 2018 Feb 4. Curr Opin Struct Biol. 2018. PMID: 29414516 Free PMC article. Review.
-
The small subunit processome in ribosome biogenesis—progress and prospects.Wiley Interdiscip Rev RNA. 2011 Jan-Feb;2(1):1-21. doi: 10.1002/wrna.57. Wiley Interdiscip Rev RNA. 2011. PMID: 21318072 Free PMC article. Review.
Cited by
-
Elucidation of the assembly events required for the recruitment of Utp20, Imp4 and Bms1 onto nascent pre-ribosomes.Nucleic Acids Res. 2011 Oct;39(18):8105-21. doi: 10.1093/nar/gkr508. Epub 2011 Jun 30. Nucleic Acids Res. 2011. PMID: 21724601 Free PMC article.
-
Ribosome Biogenesis in Plants: From Functional 45S Ribosomal DNA Organization to Ribosome Assembly Factors.Plant Cell. 2019 Sep;31(9):1945-1967. doi: 10.1105/tpc.18.00874. Epub 2019 Jun 25. Plant Cell. 2019. PMID: 31239391 Free PMC article. Review.
-
The rRNA methyltransferase Bud23 shows functional interaction with components of the SSU processome and RNase MRP.RNA. 2013 Jun;19(6):828-40. doi: 10.1261/rna.037671.112. Epub 2013 Apr 19. RNA. 2013. PMID: 23604635 Free PMC article.
-
Subcellular localization of nucleolar protein 14 and its proliferative function mediated by miR-17-5p and E2F4 in pancreatic cancer.Aging (Albany NY). 2023 Jul 27;15(14):7308-7323. doi: 10.18632/aging.204915. Epub 2023 Jul 27. Aging (Albany NY). 2023. PMID: 37506248 Free PMC article.
-
The complete structure of the small-subunit processome.Nat Struct Mol Biol. 2017 Nov;24(11):944-953. doi: 10.1038/nsmb.3472. Epub 2017 Sep 25. Nat Struct Mol Biol. 2017. PMID: 28945246
References
-
- Tschochner H, Hurt E. Pre-ribosomes on the road from the nucleolus to the cytoplasm. Trends Cell Biol. 2003;13:255–263. - PubMed
-
- Venema J, Tollervey D. Ribosome synthesis in Saccharomyces cerevisiae. Annu Rev Genet. 1999;33:261–311. - PubMed
-
- Osheim YN, French SL, Keck KM, Champion EA, Spasov K, et al. Pre-18S ribosomal RNA is structurally compacted into the SSU processome prior to being cleaved from nascent transcripts in Saccharomyces cerevisiae. Mol Cell. 2004;16:943–954. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
