

Evolution of DNA ligases of nucleo-cytoplasmic large DNA viruses of eukaryotes: a case of hidden complexity
- PMID: 20021668
- PMCID: PMC2806865
- DOI: 10.1186/1745-6150-4-51
Evolution of DNA ligases of nucleo-cytoplasmic large DNA viruses of eukaryotes: a case of hidden complexity
Abstract
Background: Eukaryotic Nucleo-Cytoplasmic Large DNA Viruses (NCLDV) encode most if not all of the enzymes involved in their DNA replication. It has been inferred that genes for these enzymes were already present in the last common ancestor of the NCLDV. However, the details of the evolution of these genes that bear on the complexity of the putative ancestral NCLDV and on the evolutionary relationships between viruses and their hosts are not well understood.
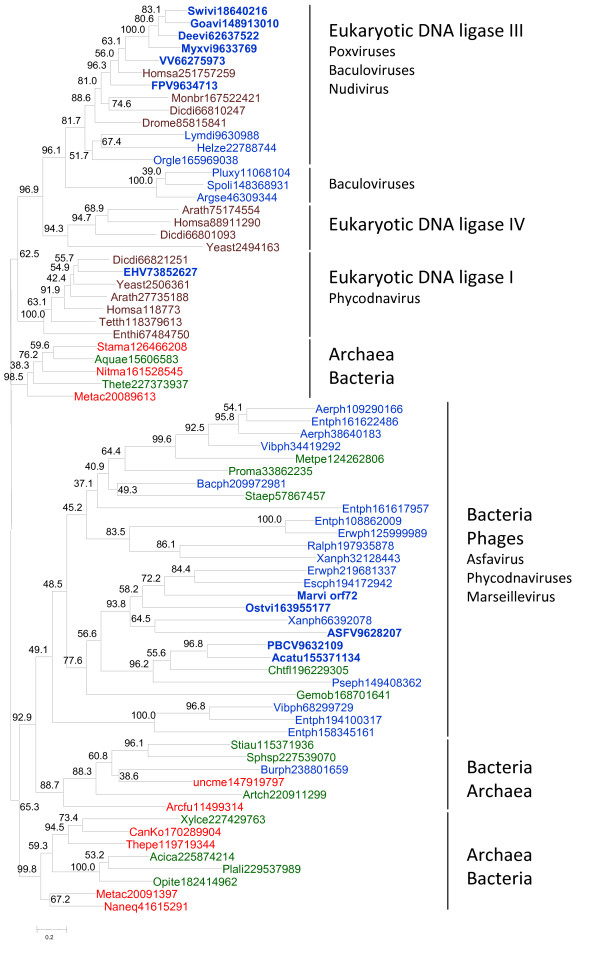
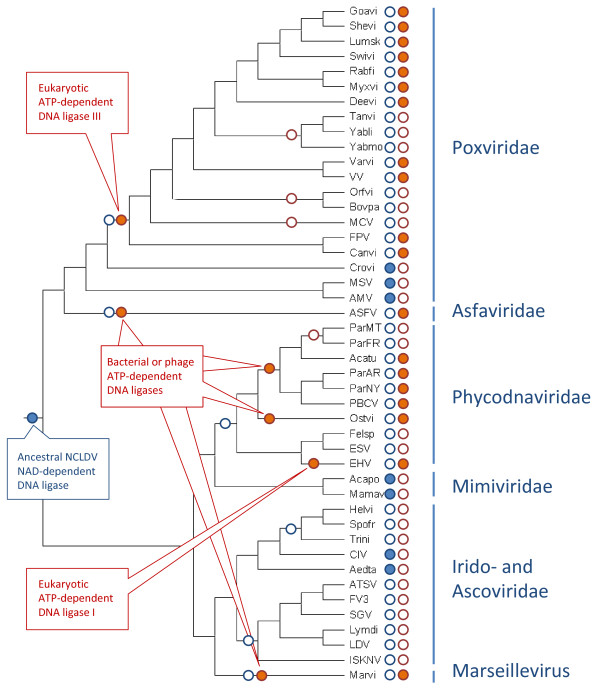
Results: Phylogenetic analysis of the ATP-dependent and NAD-dependent DNA ligases encoded by the NCLDV reveals an unexpectedly complex evolutionary history. The NAD-dependent ligases are encoded only by a minority of NCLDV (including mimiviruses, some iridoviruses and entomopoxviruses) but phylogenetic analysis clearly indicated that all viral NAD-dependent ligases are monophyletic. Combined with the topology of the NCLDV tree derived by consensus of trees for universally conserved genes suggests that this enzyme was represented in the ancestral NCLDV. Phylogenetic analysis of ATP-dependent ligases that are encoded by chordopoxviruses, most of the phycodnaviruses and Marseillevirus failed to demonstrate monophyly and instead revealed an unexpectedly complex evolutionary trajectory. The ligases of the majority of phycodnaviruses and Marseillevirus seem to have evolved from bacteriophage or bacterial homologs; the ligase of one phycodnavirus, Emiliana huxlei virus, belongs to the eukaryotic DNA ligase I branch; and ligases of chordopoxviruses unequivocally cluster with eukaryotic DNA ligase III.
Conclusions: Examination of phyletic patterns and phylogenetic analysis of DNA ligases of the NCLDV suggest that the common ancestor of the extant NCLDV encoded an NAD-dependent ligase that most likely was acquired from a bacteriophage at the early stages of evolution of eukaryotes. By contrast, ATP-dependent ligases from different prokaryotic and eukaryotic sources displaced the ancestral NAD-dependent ligase at different stages of subsequent evolution. These findings emphasize complex routes of viral evolution that become apparent through detailed phylogenomic analysis but not necessarily in reconstructions based on phyletic patterns of genes.
Reviewers: This article was reviewed by: Patrick Forterre, George V. Shpakovski, and Igor B. Zhulin.
Figures

References
-
- Boyer M, Yutin N, Pagnier I, Barrassi L, Fournous G, Espinosa M, Robert C, Azza A, Sun S, Rossmann MG, Suzan-Monti M, La Scola B, Koonin EV, Raoult D. Giant Marseillevirus highlights the role of amoebae as a melting pot in emergence of chimaeric microorganisms. Proc Natl Acad Sci USA. 2009. in press . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
