

How antibodies fold
- PMID: 20022755
- PMCID: PMC4716677
- DOI: 10.1016/j.tibs.2009.11.005
How antibodies fold
Abstract
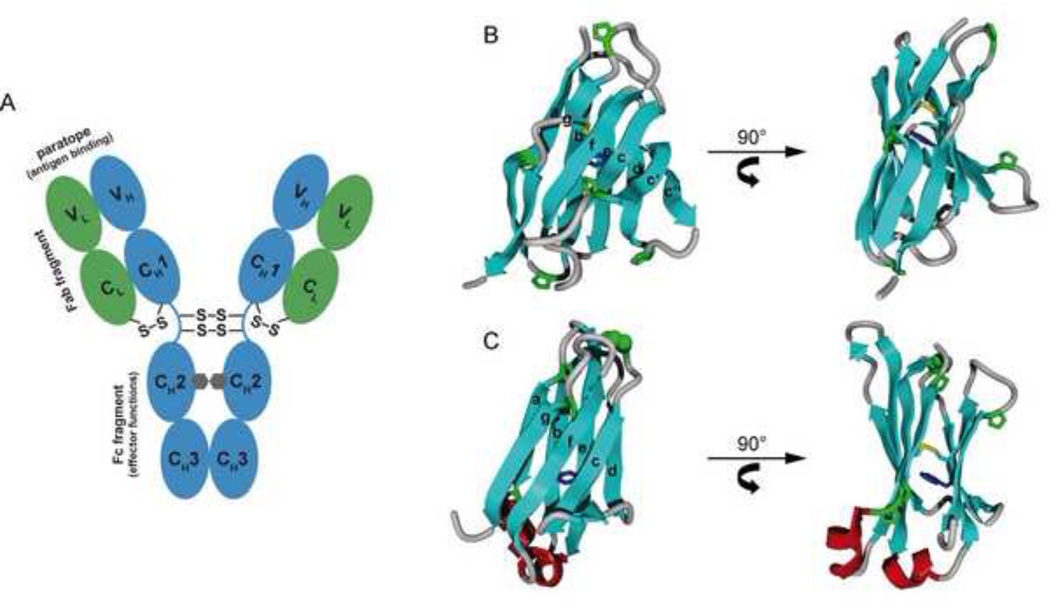
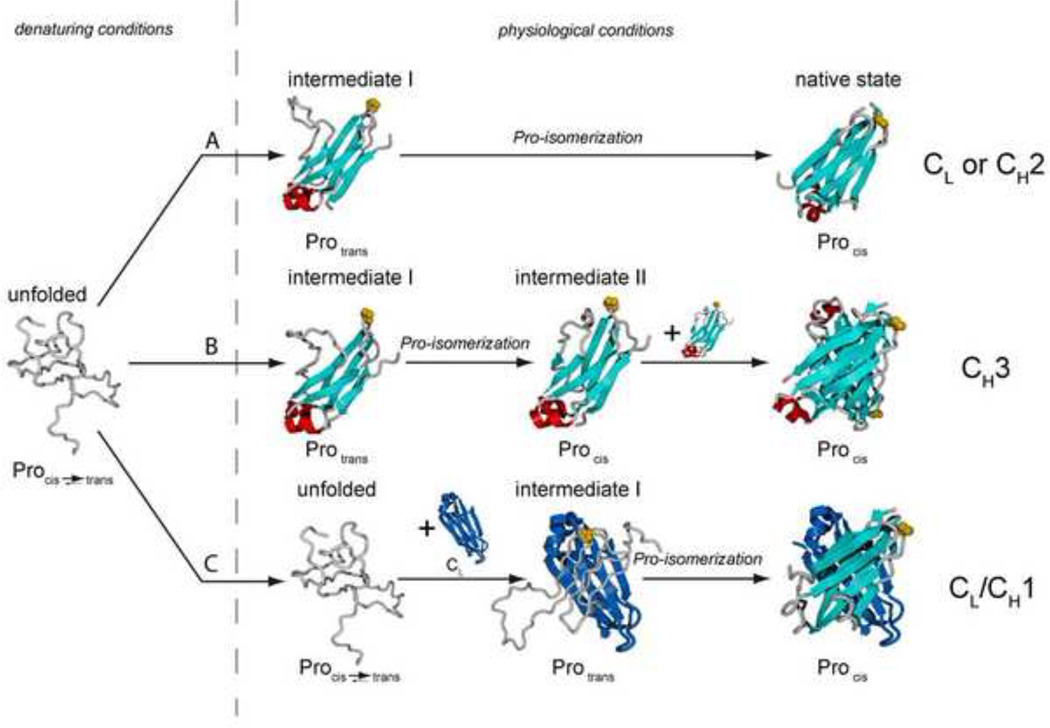
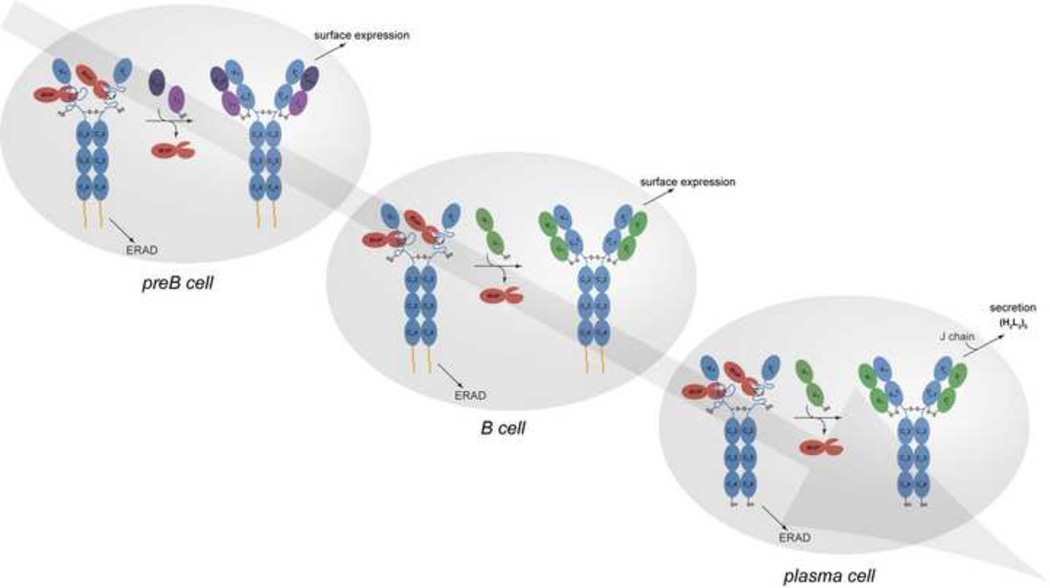
B cells use unconventional strategies for the production of a seemingly unlimited number of antibodies from a very limited amount of DNA. These methods dramatically increase the likelihood of producing proteins that cannot fold or assemble appropriately. B cells are therefore particularly dependent on 'quality control' mechanisms to oversee antibody production. Recent in vitro experiments demonstrate that Ig domains have evolved diverse folding strategies ranging from robust spontaneous folding to intrinsically disordered domains that require assembly with their partner domains to fold; in vivo experiments reveal that these different folding characteristics form the basis for cellular checkpoints in Ig transport. Taken together, these reports provide a detailed understanding of how B cells monitor and ensure the functional fidelity of Ig proteins.
2009 Elsevier Ltd. All rights reserved.
Figures

Comment in
-
Immunoglobulin transport in the absence of light chains.Trends Biochem Sci. 2010 Nov;35(11):593; author reply 594. doi: 10.1016/j.tibs.2010.07.010. Epub 2010 Aug 20. Trends Biochem Sci. 2010. PMID: 20728360 No abstract available.
References
-
- Murphy KM, et al. Janeway's Immunobiology. Garland Science; 2008.
-
- Early P, et al. An immunoglobulin heavy chain variable region gene is generated from three segments of DNA: VH D and JH. Cell. 1980;19:981–992. - PubMed
-
- Calame K, et al. Mouse Cmu heavy chain immunoglobulin gene segment contains three intervening sequences separating domains. Nature. 1980;284:452–455. - PubMed
-
- Alt FW, et al. Development of the primary antibody repertoire. Science. 1987;238:1079–1087. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
