

mTOR Ser-2481 autophosphorylation monitors mTORC-specific catalytic activity and clarifies rapamycin mechanism of action
- PMID: 20022946
- PMCID: PMC2832937
- DOI: 10.1074/jbc.M109.096222
mTOR Ser-2481 autophosphorylation monitors mTORC-specific catalytic activity and clarifies rapamycin mechanism of action
Abstract
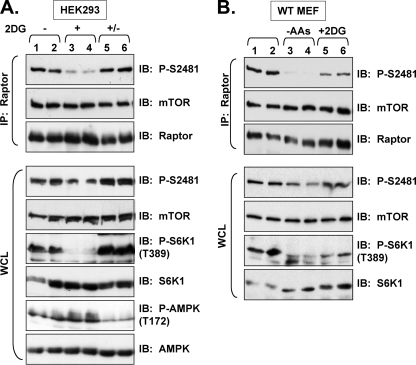
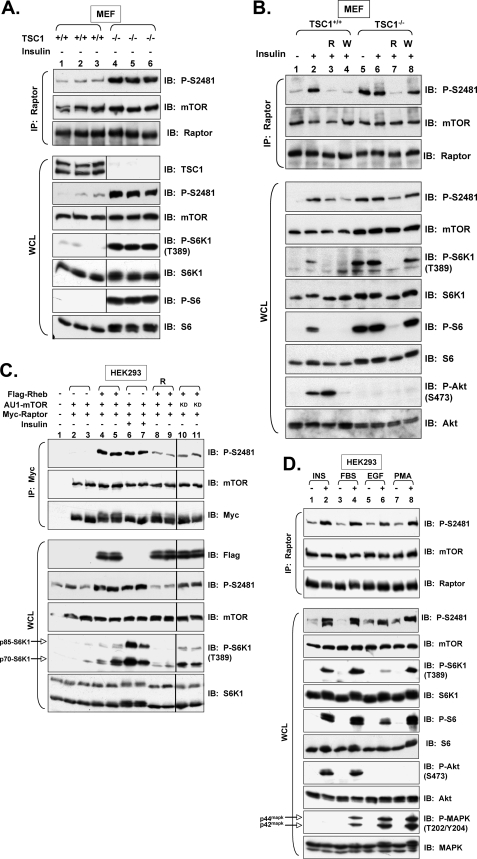
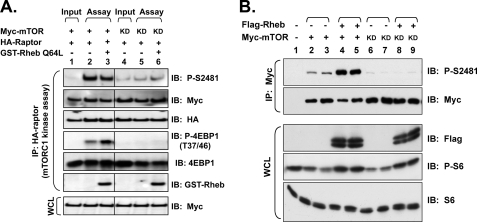
The mammalian target of rapamycin (mTOR) Ser/Thr kinase signals in at least two multiprotein complexes distinguished by their different partners and sensitivities to rapamycin. Acute rapamycin inhibits signaling by mTOR complex 1 (mTORC1) but not mTOR complex 2 (mTORC2), which both promote cell growth, proliferation, and survival. Although mTORC2 regulation remains poorly defined, diverse cellular mitogens activate mTORC1 signaling in a manner that requires sufficient levels of amino acids and cellular energy. Before the identification of distinct mTOR complexes, mTOR was reported to autophosphorylate on Ser-2481 in vivo in a rapamycin- and amino acid-insensitive manner. These results suggested that modulation of mTOR intrinsic catalytic activity does not universally underlie mTOR regulation. Here we re-examine the regulation of mTOR Ser-2481 autophosphorylation (Ser(P)-2481) in vivo by studying mTORC-specific Ser(P)-2481 in mTORC1 and mTORC2, with a primary focus on mTORC1. In contrast to previous work, we find that acute rapamycin and amino acid withdrawal markedly attenuate mTORC1-associated mTOR Ser(P)-2481 in cycling cells. Although insulin stimulates both mTORC1- and mTORC2-associated mTOR Ser(P)-2481 in a phosphatidylinositol 3-kinase-dependent manner, rapamycin acutely inhibits insulin-stimulated mTOR Ser(P)-2481 in mTORC1 but not mTORC2. By interrogating diverse mTORC1 regulatory input, we find that without exception mTORC1-activating signals promote, whereas mTORC1-inhibitory signals decrease mTORC1-associated mTOR Ser(P)-2481. These data suggest that mTORC1- and likely mTORC2-associated mTOR Ser-2481 autophosphorylation directly monitors intrinsic mTORC-specific catalytic activity and reveal that rapamycin inhibits mTORC1 signaling in vivo by reducing mTORC1 catalytic activity.
Figures
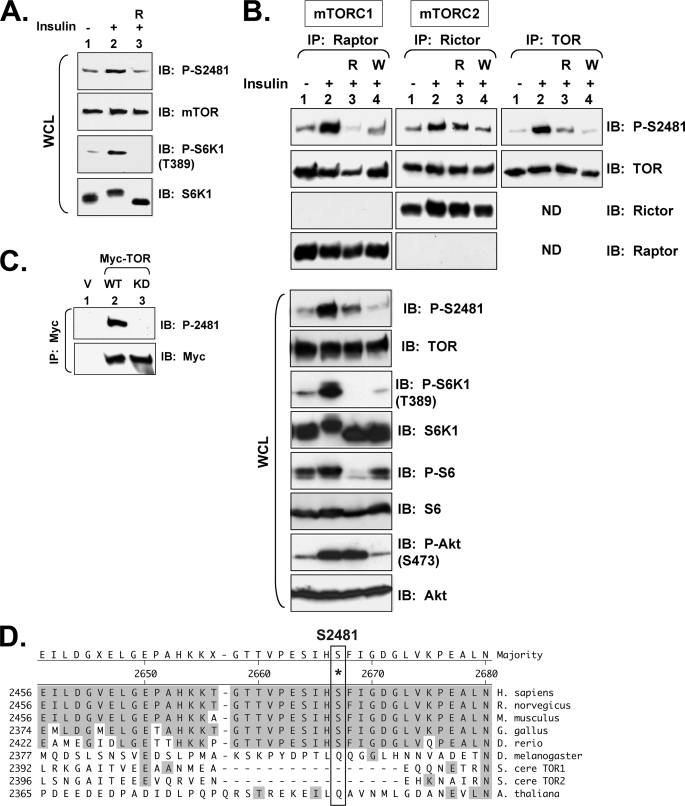
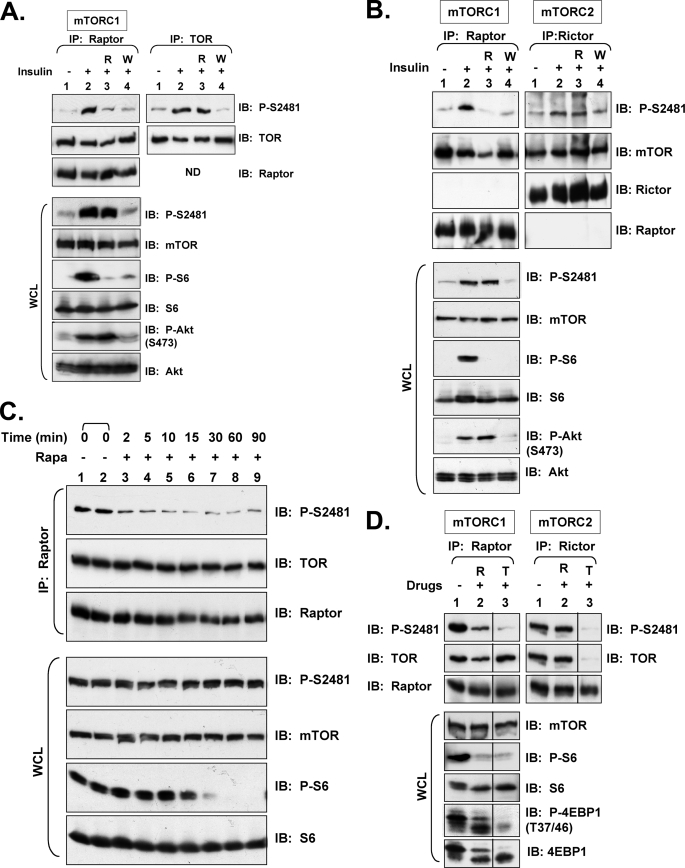
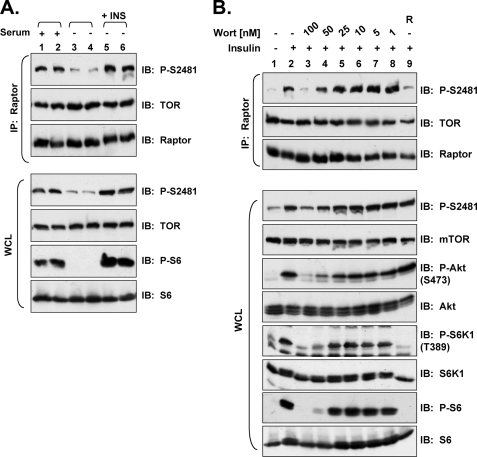
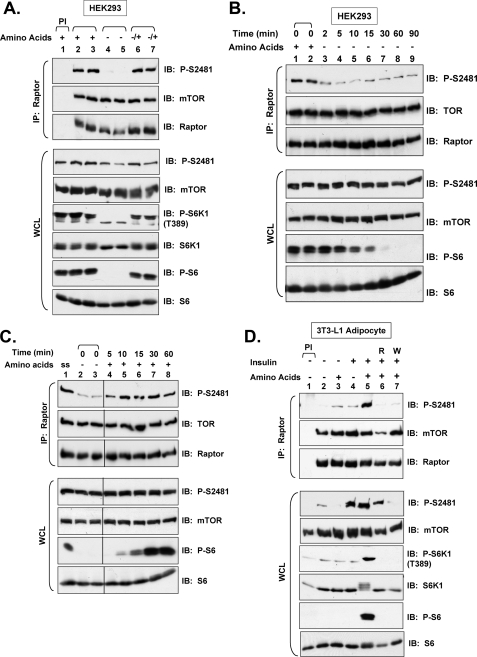
References
-
- Dunlop E. A., Tee A. R. (2009) Cell. Signal. 21, 827–835 - PubMed
-
- Fingar D. C., Blenis J. (2004) Oncogene 23, 3151–3171 - PubMed
-
- Jacinto E., Lorberg A. (2008) Biochem. J. 410, 19–37 - PubMed
-
- Bhaskar P. T., Hay N. (2007) Dev. Cell 12, 487–502 - PubMed
-
- Guertin D. A., Sabatini D. M. (2009) Sci. Signal. 2, pe24. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
