

Characterization of CamH from Methanosarcina thermophila, founding member of a subclass of the {gamma} class of carbonic anhydrases
- PMID: 20023030
- PMCID: PMC2820857
- DOI: 10.1128/JB.01164-09
Characterization of CamH from Methanosarcina thermophila, founding member of a subclass of the {gamma} class of carbonic anhydrases
Abstract
The homotrimeric enzyme Mt-Cam from Methanosarcina thermophila is the archetype of the gamma class of carbonic anhydrases. A search of databases queried with Mt-Cam revealed that a majority of the homologs comprise a putative subclass (CamH) in which there is major conservation of all of the residues essential for the archetype Mt-Cam except Glu62 and an acidic loop containing the essential proton shuttle residue Glu84. The CamH homolog from M. thermophila (Mt-CamH) was overproduced in Escherichia coli and characterized to validate its activity and initiate an investigation of the CamH subclass. The Mt-CamH homotrimer purified from E. coli cultured with supplemental zinc (Zn-Mt-CamH) contained 0.71 zinc and 0.15 iron per monomer and had k(cat) and k(cat)/K(m) values that were substantially lower than those for the zinc form of Mt-Cam (Zn-Mt-Cam). Mt-CamH purified from E. coli cultured with supplemental iron (Fe-Mt-CamH) was also a trimer containing 0.15 iron per monomer and only a trace amount of zinc and had an effective k(cat) (k(cat)(eff)) value normalized for iron that was 6-fold less than that for the iron form of Mt-Cam, whereas the k(cat)/K(m)(eff) was similar to that for Fe-Mt-Cam. Addition of 50 mM imidazole to the assay buffer increased the k(cat)(eff) of Fe-Mt-CamH more than 4-fold. Fe-Mt-CamH lost activity when it was exposed to air or 3% H(2)O(2), which supports the hypothesis that Fe(2+) has a role in the active site. The k(cat) for Fe-Mt-CamH was dependent on the concentration of buffer in a way that indicates that it acts as a second substrate in a "ping-pong" mechanism accepting a proton. The k(cat)/K(m) was not dependent on the buffer, consistent with the mechanism for all carbonic anhydrases in which the interconversion of CO(2) and HCO(3)(-) is separate from intermolecular proton transfer.
Figures

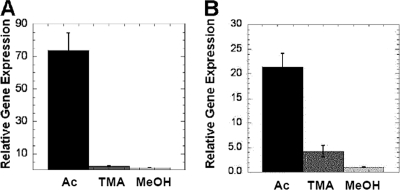
Similar articles
-
Kinetic and spectroscopic characterization of the gamma-carbonic anhydrase from the methanoarchaeon Methanosarcina thermophila.Biochemistry. 1999 Oct 5;38(40):13119-28. doi: 10.1021/bi9828876. Biochemistry. 1999. PMID: 10529183
-
A structure-function study of a proton transport pathway in the gamma-class carbonic anhydrase from Methanosarcina thermophila.Biochemistry. 2000 Aug 8;39(31):9232-40. doi: 10.1021/bi0001877. Biochemistry. 2000. PMID: 10924116
-
A closer look at the active site of gamma-class carbonic anhydrases: high-resolution crystallographic studies of the carbonic anhydrase from Methanosarcina thermophila.Biochemistry. 2000 Aug 8;39(31):9222-31. doi: 10.1021/bi000204s. Biochemistry. 2000. PMID: 10924115
-
The gamma class of carbonic anhydrases.Biochim Biophys Acta. 2010 Feb;1804(2):374-81. doi: 10.1016/j.bbapap.2009.08.026. Epub 2009 Sep 10. Biochim Biophys Acta. 2010. PMID: 19747990 Free PMC article. Review.
-
Prokaryotic carbonic anhydrases.FEMS Microbiol Rev. 2000 Oct;24(4):335-66. doi: 10.1111/j.1574-6976.2000.tb00546.x. FEMS Microbiol Rev. 2000. PMID: 10978542 Review.
Cited by
-
Molecular structure of the Brucella abortus metalloprotein RicA, a Rab2-binding virulence effector.Biochemistry. 2013 Dec 17;52(50):9020-8. doi: 10.1021/bi401373r. Epub 2013 Nov 22. Biochemistry. 2013. PMID: 24251537 Free PMC article.
-
Identification and characterization of a carboxysomal γ-carbonic anhydrase from the cyanobacterium Nostoc sp. PCC 7120.Photosynth Res. 2014 Sep;121(2-3):135-50. doi: 10.1007/s11120-014-0018-4. Epub 2014 Jun 8. Photosynth Res. 2014. PMID: 24907906
-
Crystallography and Its Impact on Carbonic Anhydrase Research.Int J Med Chem. 2018 Sep 13;2018:9419521. doi: 10.1155/2018/9419521. eCollection 2018. Int J Med Chem. 2018. PMID: 30302289 Free PMC article. Review.
-
Expression of seven carbonic anhydrases in red alga Gracilariopsis chorda and their subcellular localization in a heterologous system, Arabidopsis thaliana.Plant Cell Rep. 2019 Feb;38(2):147-159. doi: 10.1007/s00299-018-2356-8. Epub 2018 Nov 16. Plant Cell Rep. 2019. PMID: 30446790
-
Model Organisms To Study Methanogenesis, a Uniquely Archaeal Metabolism.J Bacteriol. 2023 Aug 24;205(8):e0011523. doi: 10.1128/jb.00115-23. Epub 2023 Jul 17. J Bacteriol. 2023. PMID: 37458589 Free PMC article. Review.
References
-
- Alber, B. E., C. M. Colangelo, J. Dong, C. M. V. Stålhandske, T. T. Baird, C. Tu, C. A. Fierke, D. N. Silverman, R. A. Scott, and J. G. Ferry. 1999. Kinetic and spectroscopic characterization of the gamma-carbonic anhydrase from the methanoarchaeon Methanosarcina thermophila. Biochemistry 38:13119-13128. - PubMed
-
- An, H., C. Tu, D. Duda, I. Montanez-Clemente, K. Math, P. J. Laipis, R. McKenna, and D. N. Silverman. 2002. Chemical rescue in catalysis by human carbonic anhydrases II and III. Biochemistry 41:3235-3242. - PubMed
-
- Dereeper, A., V. Guignon, G. Blanc, S. Audic, S. Buffet, F. Chevenet, J. F. Dufayard, S. Guindon, V. Lefort, M. Lescot, J. M. Claverie, and O. Gascuel. 2008. Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36:W465-W469. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
