

Tryptophan-derived metabolites are required for antifungal defense in the Arabidopsis mlo2 mutant
- PMID: 20023151
- PMCID: PMC2832281
- DOI: 10.1104/pp.109.147660
Tryptophan-derived metabolites are required for antifungal defense in the Arabidopsis mlo2 mutant
Abstract
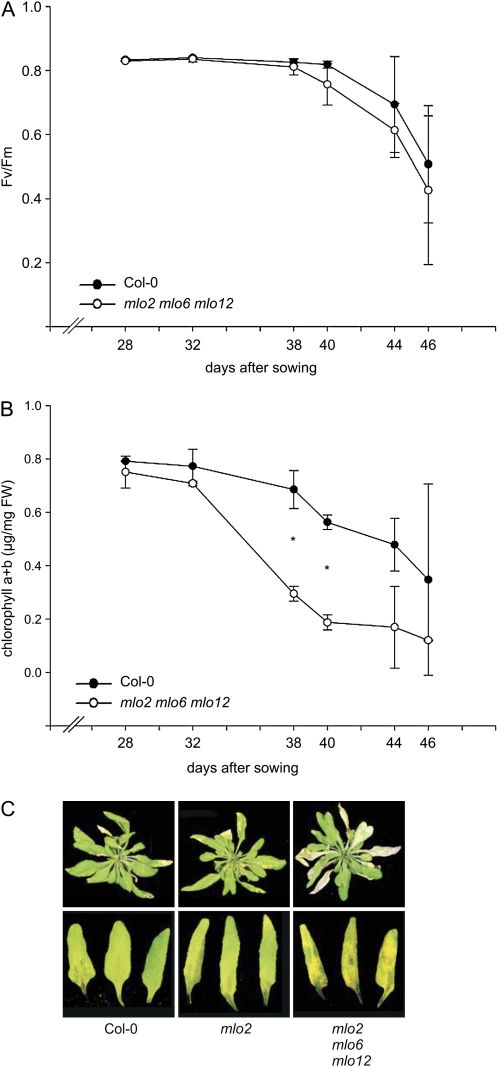
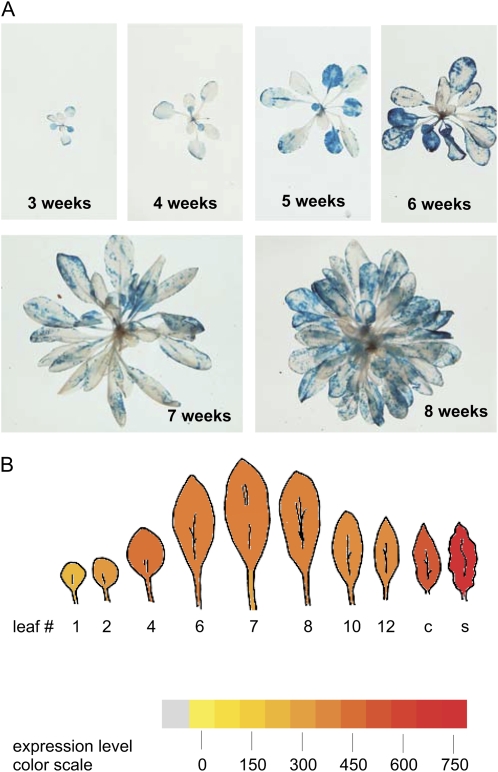
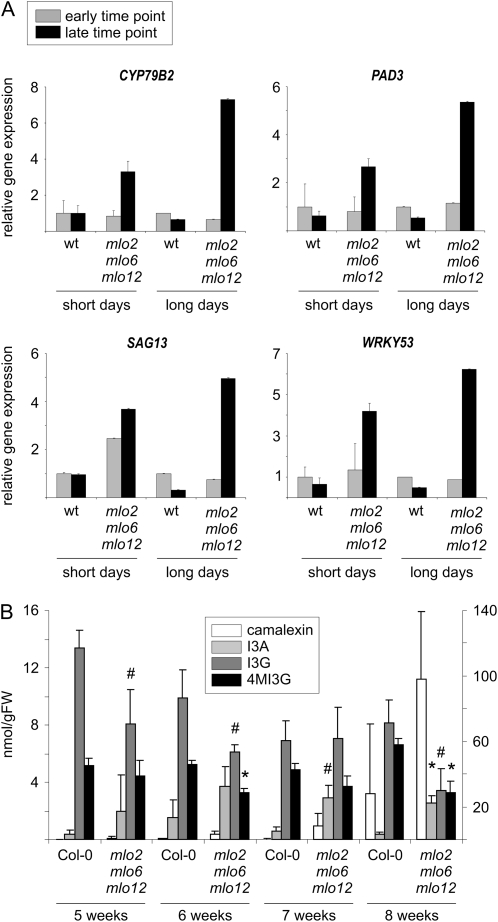
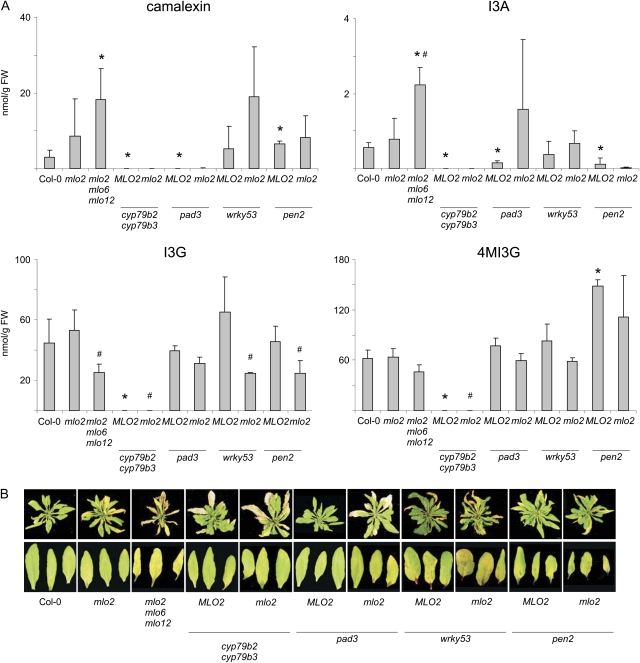
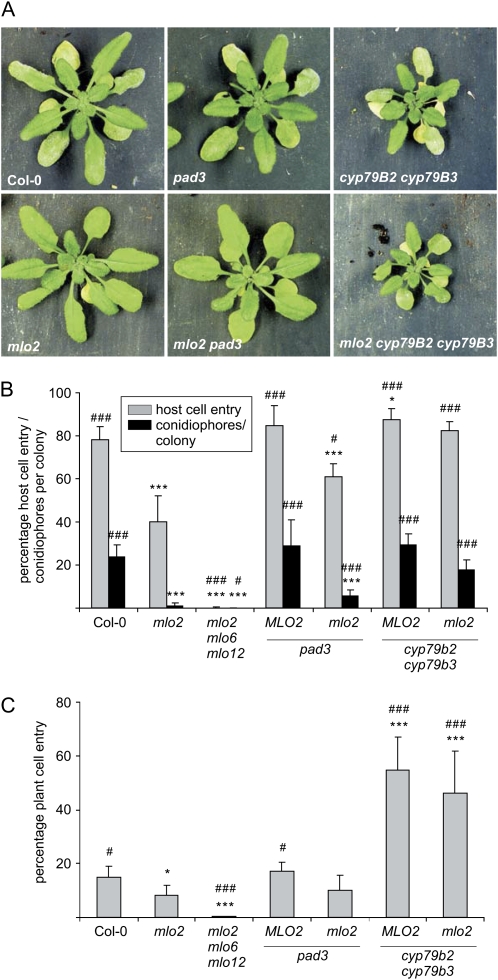
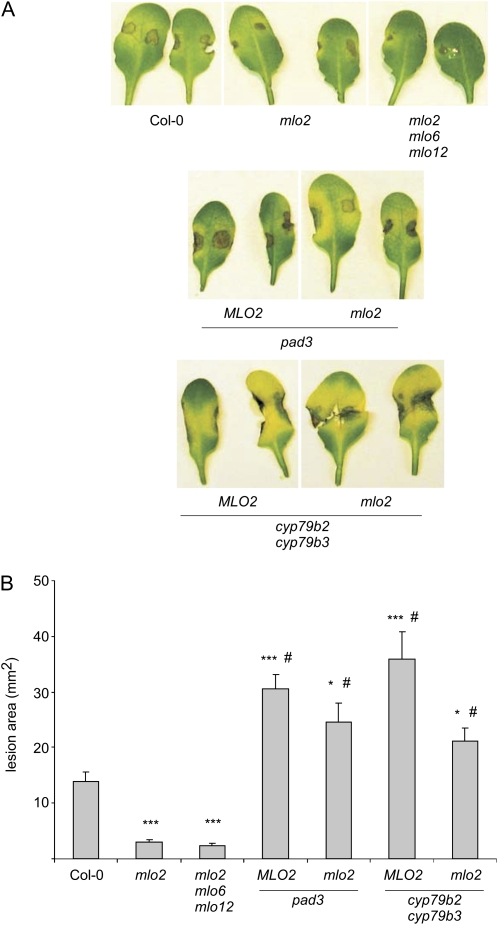
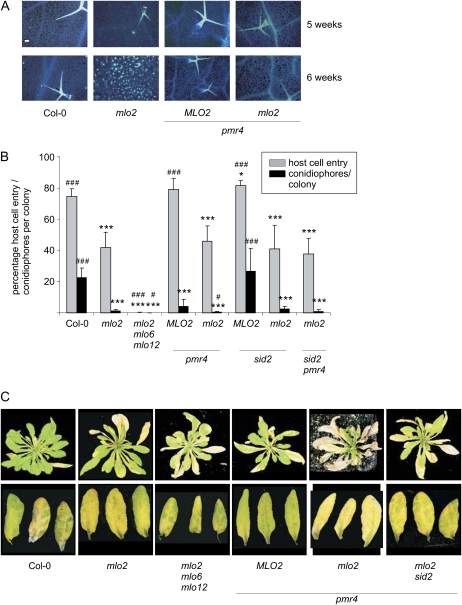
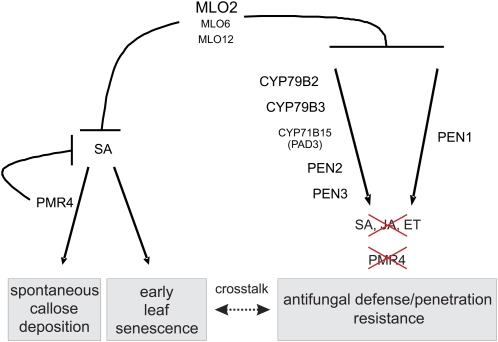
Arabidopsis (Arabidopsis thaliana) genes MILDEW RESISTANCE LOCUS O2 (MLO2), MLO6, and MLO12 exhibit unequal genetic redundancy with respect to the modulation of defense responses against powdery mildew fungi and the control of developmental phenotypes such as premature leaf decay. We show that early chlorosis and necrosis of rosette leaves in mlo2 mlo6 mlo12 mutants reflects an authentic but untimely leaf senescence program. Comparative transcriptional profiling revealed that transcripts of several genes encoding tryptophan biosynthetic and metabolic enzymes hyperaccumulate during vegetative development in the mlo2 mlo6 mlo12 mutant. Elevated expression levels of these genes correlate with altered steady-state levels of several indolic metabolites, including the phytoalexin camalexin and indolic glucosinolates, during development in the mlo2 single mutant and the mlo2 mlo6 mlo12 triple mutant. Results of genetic epistasis analysis suggest a decisive role for indolic metabolites in mlo2-conditioned antifungal defense against both biotrophic powdery mildews and a camalexin-sensitive strain of the necrotrophic fungus Botrytis cinerea. The wound- and pathogen-responsive callose synthase POWDERY MILDEW RESISTANCE4/GLUCAN SYNTHASE-LIKE5 was found to be responsible for the spontaneous callose deposits in mlo2 mutant plants but dispensable for mlo2-conditioned penetration resistance. Our data strengthen the notion that powdery mildew resistance of mlo2 genotypes is based on the same defense execution machinery as innate antifungal immune responses that restrict the invasion of nonadapted fungal pathogens.
Figures
Similar articles
-
The powdery mildew-resistant Arabidopsis mlo2 mlo6 mlo12 triple mutant displays altered infection phenotypes with diverse types of phytopathogens.Sci Rep. 2017 Aug 24;7(1):9319. doi: 10.1038/s41598-017-07188-7. Sci Rep. 2017. PMID: 28839137 Free PMC article.
-
Key Components of Different Plant Defense Pathways Are Dispensable for Powdery Mildew Resistance of the Arabidopsis mlo2 mlo6 mlo12 Triple Mutant.Front Plant Sci. 2017 Jun 19;8:1006. doi: 10.3389/fpls.2017.01006. eCollection 2017. Front Plant Sci. 2017. PMID: 28674541 Free PMC article.
-
Arabidopsis MLO2 is a negative regulator of sensitivity to extracellular reactive oxygen species.Plant Cell Environ. 2018 Apr;41(4):782-796. doi: 10.1111/pce.13144. Epub 2018 Feb 22. Plant Cell Environ. 2018. PMID: 29333607
-
Linking phytochrome to plant immunity: low red : far-red ratios increase Arabidopsis susceptibility to Botrytis cinerea by reducing the biosynthesis of indolic glucosinolates and camalexin.New Phytol. 2014 Oct;204(2):342-54. doi: 10.1111/nph.13032. New Phytol. 2014. PMID: 25236170
-
A critical role for Arabidopsis MILDEW RESISTANCE LOCUS O2 in systemic acquired resistance.Plant J. 2018 Jun;94(6):1064-1082. doi: 10.1111/tpj.13920. Epub 2018 May 10. Plant J. 2018. PMID: 29660188
Cited by
-
The differential expression patterns of paralogs in response to stresses indicate expression and sequence divergences.BMC Plant Biol. 2020 Jun 16;20(1):277. doi: 10.1186/s12870-020-02460-x. BMC Plant Biol. 2020. PMID: 32546126 Free PMC article.
-
Cucumber Mildew Resistance Locus O Interacts with Calmodulin and Regulates Plant Cell Death Associated with Plant Immunity.Int J Mol Sci. 2019 Jun 19;20(12):2995. doi: 10.3390/ijms20122995. Int J Mol Sci. 2019. PMID: 31248151 Free PMC article.
-
Callose-mediated resistance to pathogenic intruders in plant defense-related papillae.Front Plant Sci. 2014 Apr 28;5:168. doi: 10.3389/fpls.2014.00168. eCollection 2014. Front Plant Sci. 2014. PMID: 24808903 Free PMC article.
-
Chemical suppressors of mlo-mediated powdery mildew resistance.Biosci Rep. 2017 Dec 12;37(6):BSR20171389. doi: 10.1042/BSR20171389. Print 2017 Dec 22. Biosci Rep. 2017. PMID: 29127104 Free PMC article.
-
The powdery mildew-resistant Arabidopsis mlo2 mlo6 mlo12 triple mutant displays altered infection phenotypes with diverse types of phytopathogens.Sci Rep. 2017 Aug 24;7(1):9319. doi: 10.1038/s41598-017-07188-7. Sci Rep. 2017. PMID: 28839137 Free PMC article.
References
-
- Adam L, Somerville SC. (1996) Genetic characterization of five powdery mildew disease resistance loci in Arabidopsis thaliana. Plant J 9: 341–356 - PubMed
-
- Aist JR. (1976) Papillae and related wound plugs of plant cells. Annu Rev Phytopathol 14: 145–163
-
- Bayles CJ, Ghemawat MS, Aist JR. (1990) Inhibition by 2-deoxy-D-glucose of callose formation, papilla deposition, and resistance to powdery mildew in an ml-o barley mutant. Physiol Mol Plant Pathol 36: 63–72
-
- Bednarek P, Pislewska-Bednarek M, Svatos A, Schneider B, Doubsky J, Mansurova M, Humphry M, Consonni C, Panstruga R, Sanchez-Vallet A, et al. (2009) A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323: 101–106 - PubMed
-
- Beißbarth T, Speed TP. (2004) GOstat: find statistically overrepresented Gene Ontologies within a group of genes. Bioinformatics 20: 1464–1465 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
