

Review
doi: 10.1242/dev.036160.
Dissecting the regulatory switches of development: lessons from enhancer evolution in Drosophila
Affiliations
- PMID: 20023155
- PMCID: PMC2796927
- DOI: 10.1242/dev.036160
Item in Clipboard
Review
Dissecting the regulatory switches of development: lessons from enhancer evolution in Drosophila
Development.
2010 Jan.
Abstract
Cis-regulatory modules are non-protein-coding regions of DNA essential for the control of gene expression. One class of regulatory modules is embryonic enhancers, which drive gene expression during development as a result of transcription factor protein binding at the enhancer sequences. Recent comparative studies have begun to investigate the evolution of the sequence architecture within enhancers. These analyses are illuminating the way that developmental biologists think about enhancers by revealing their molecular mechanism of function.
Figures
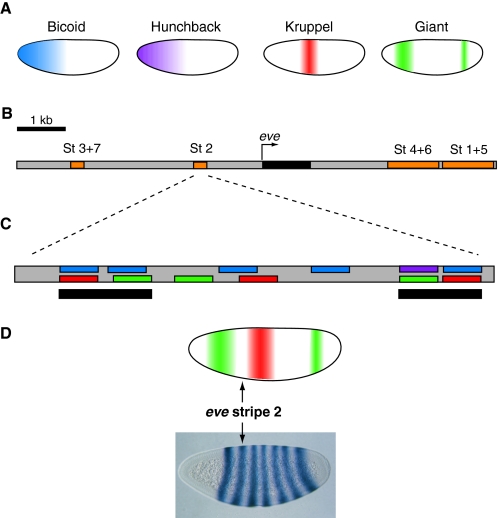
Transcription factor gradients activate even skipped expression. (A) The localization patterns of four major transcription factors (TFs) in the early Drosophila embryo are shown. Embryos are oriented anterior to the left and dorsal is up. The Bicoid (blue) and Hunchback (purple) TFs are activators of the even skipped (eve) stripe 2 enhancer, whereas the Kruppel (red) and Giant (green) TFs are repressors. Both activators are broadly expressed in the anterior half of the embryo, but repressor expression is more restricted. (B) Map of the eve genomic locus, including the embryonic enhancers (orange) responsible for eve (black) expression. Each enhancer drives eve expression in one or two developing parasegments of the embryo. Together, the enhancers drive expression in seven stripes (St 1-7) in odd-numbered parasegments. (C) The minimal eve stripe 2 enhancer (St 2) of D. melanogaster contains twelve transcription factor binding sites. The locations of color-coded binding sites for the activators and repressors shown in A are indicated. Two clusters of binding sites (black bars) at each end of the minimal stripe 2 module are thought to be particularly important for functional activity. (D) Sharp borders of eve stripe 2-directed expression are established by high concentrations of Giant (green) at the anterior boundary and Kruppel (red) at the posterior boundary.
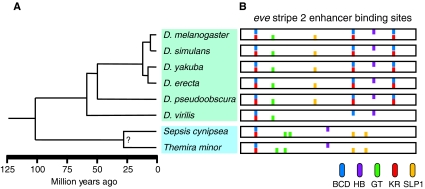
Phylogenetic relationship and eve stripe 2 enhancer architecture of Drosophila and Sepsid species. (A) The Drosophila (green box) diverged from the Sepsid family (blue box) approximately 100 million years ago (Mya). The distantly related Drosophila species, D. melanogaster and D. virilis, diverged 60 Mya (Tamura et al., 2004). (B) The organization of a subset of bioinformatically predicted binding sites for the transcription factors Bicoid (BCD, blue), Hunchback (HB, purple), Giant (GT, green), Kruppel (KR, red) and Sloppy paired 1 (SLP1, yellow) within the minimal eve stripe 2 enhancer [summarized from Hare (Hare et al., 2008a)] are shown for eight species: D. melanogaster, D. simulans, D. yakuba, D. erecta, D. pseudoobscura, D. virilis, Sepsis cynipsea and Themira minor.
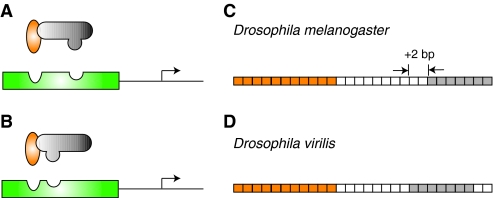
Changing regulatory environments can drive compensatory changes in enhancer organization. In (A) D. melanogaster and (B) D. virilis, the Dorsal (orange) and Twist (gray) activators cooperatively bind to neurogenic ectoderm enhancer (NEE) sequences (green) to upregulate the transcription of target genes (arrow). This interaction is dependent upon compatibility between the transcription factors and their respective binding sites. In the ~60 million years of evolution separating D. virilis and D. melanogaster (Tamura et al., 2004), protein sequences have diverged only slightly, but enough to affect the DNA-binding domains of the proteins. As a result, in the vnd NEEs of (C) D. melanogaster and (D) D. virilis, the spacing between a conserved Dorsal binding site (orange boxes) and a conserved Twist binding site (gray boxes) is different. The intervening sequence between the binding sites is only 37.5% conserved, but there is also a deletion of two nucleotides (white boxes) in the D. virilis NEE. Even small changes in transcription factor protein domains between different species can drive the architectural reorganization of DNA binding sites. In this case, a decrease in the spacing between sites preserves enhancer functional activity.
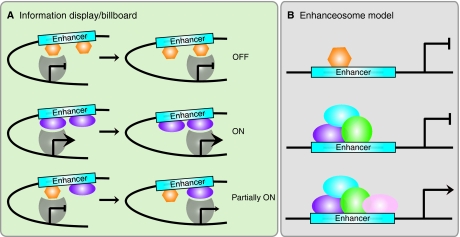
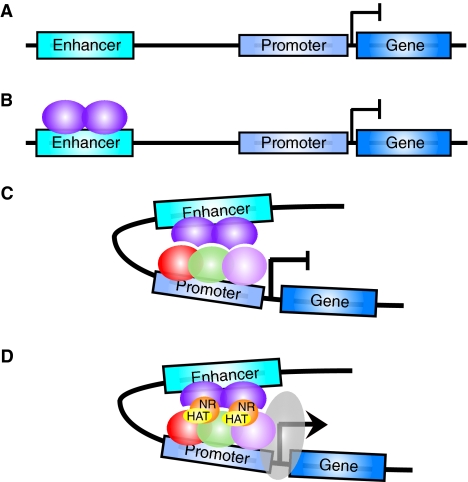
Information display/billboard and enhanceosome models of cis-regulatory module function. (A) In the information display/billboard model, the transcriptional machinery (gray shape) samples the regulatory landscape found at an enhancer module (blue box) (Kulkarni and Arnosti, 2003). If the basal transcriptional machinery encounters only repressors (orange hexagons), the target gene will not be upregulated. The binding of only activators (purple ellipses) at the regulatory module will result in strong target gene expression. A combination of signals from specific DNA sites being bound by repressors and activators will lead to an intermediate (or more probably spatiotemporally restricted) level of target gene activity. An information display enhancer is tolerant of evolutionary binding site turnover because the transcriptional machinery samples only discrete regions of the enhancer and the output signal will not necessarily differ if transcription factor binding sites are located in different regions within the enhancer sequence. (B) In an enhanceosome, the enhancer can only function when a large regulatory protein complex has assembled (purple, blue, green and pink ellipses). If a repressor (orange hexagon) occludes binding sites, or if one of the components of the complex is absent, the enhancer will not drive target gene expression. An enhanceosome will not tolerate binding site turnover, as the protein complex is extremely stereospecific.
Similar articles
-
Genome-scale functional characterization of Drosophila developmental enhancers in vivo.Nature. 2014 Aug 7;512(7512):91-5. doi: 10.1038/nature13395. Epub 2014 Jun 1. Nature. 2014. PMID: 24896182
-
Extraction of functional binding sites from unique regulatory regions: the Drosophila early developmental enhancers.Genome Res. 2002 Mar;12(3):470-81. doi: 10.1101/gr.212502. Genome Res. 2002. PMID: 11875036 Free PMC article.
-
Evolution of developmental genes: molecular microevolution of enhancer sequences at the Ubx locus in Drosophila and its impact on developmental phenotypes.Mol Biol Evol. 2004 Feb;21(2):348-63. doi: 10.1093/molbev/msh025. Epub 2003 Dec 5. Mol Biol Evol. 2004. PMID: 14660693
-
Dynamic patterning by morphogens illuminated by cis-regulatory studies.Development. 2021 Jan 20;148(2):dev196113. doi: 10.1242/dev.196113. Development. 2021. PMID: 33472851 Free PMC article. Review.
-
Tempo and mode in evolution of transcriptional regulation.PLoS Genet. 2012 Jan;8(1):e1002432. doi: 10.1371/journal.pgen.1002432. Epub 2012 Jan 19. PLoS Genet. 2012. PMID: 22291600 Free PMC article. Review.
Cited by
-
Intronic cis-regulatory modules mediate tissue-specific and microbial control of angptl4/fiaf transcription.PLoS Genet. 2012;8(3):e1002585. doi: 10.1371/journal.pgen.1002585. Epub 2012 Mar 29. PLoS Genet. 2012. PMID: 22479192 Free PMC article.
-
Color-Patterns to Architecture Conversion through Conditional Generative Adversarial Networks.Biomimetics (Basel). 2021 Feb 17;6(1):16. doi: 10.3390/biomimetics6010016. Biomimetics (Basel). 2021. PMID: 33671287 Free PMC article.
-
Role of architecture in the function and specificity of two Notch-regulated transcriptional enhancer modules.PLoS Genet. 2012 Jul;8(7):e1002796. doi: 10.1371/journal.pgen.1002796. Epub 2012 Jul 5. PLoS Genet. 2012. PMID: 22792075 Free PMC article.
-
In vivo mapping of mutagenesis sensitivity of human enhancers.Nature. 2025 Jul;643(8072):839-846. doi: 10.1038/s41586-025-09182-w. Epub 2025 Jun 18. Nature. 2025. PMID: 40533554
-
Mutagenesis Sensitivity Mapping of Human Enhancers In Vivo.bioRxiv [Preprint]. 2024 Sep 8:2024.09.06.611737. doi: 10.1101/2024.09.06.611737. bioRxiv. 2024. Update in: Nature. 2025 Jul;643(8072):839-846. doi: 10.1038/s41586-025-09182-w. PMID: 39282388 Free PMC article. Updated. Preprint.
References
-
- Andrioli L. P., Vasisht V., Theodosopoulou E., Oberstein A., Small S. (2002). Anterior repression of a Drosophila stripe enhancer requires three position-specific mechanisms. Development 129, 4931-4940 - PubMed
-
- Arnosti D. N., Barolo S., Levine M., Small S. (1996). The eve stripe 2 enhancer employs multiple modes of transcriptional synergy. Development 122, 205-214 - PubMed
-
- Banerji J., Rusconi S., Schaffner W. (1981). Expression of a beta-globin gene is enhanced by remote SV40 DNA sequences. Cell 27, 299-308 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
