

Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster
- PMID: 20023160
- PMCID: PMC2796924
- DOI: 10.1242/dev.041749
Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster
Abstract
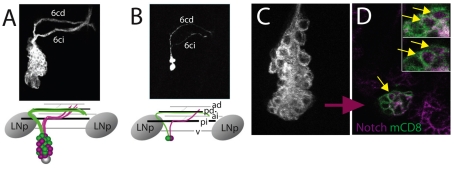
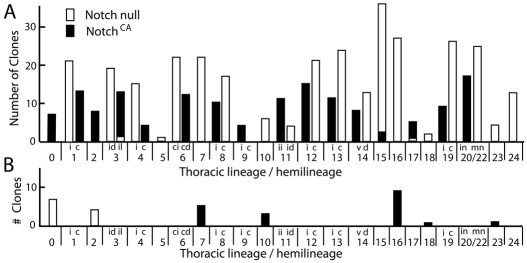
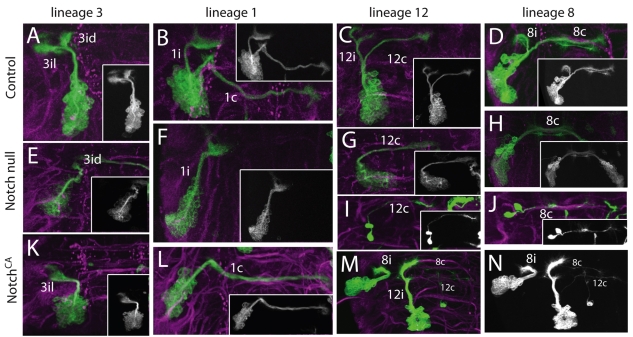
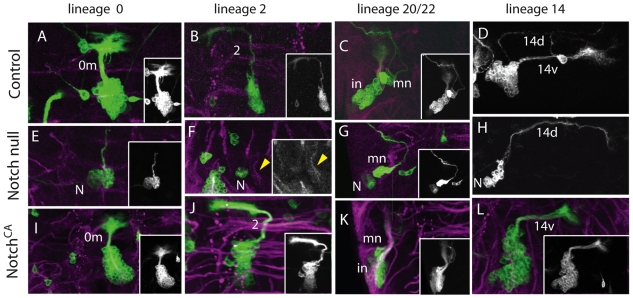
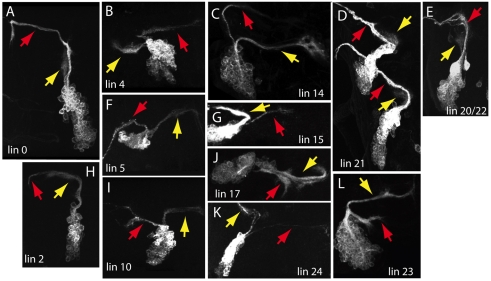
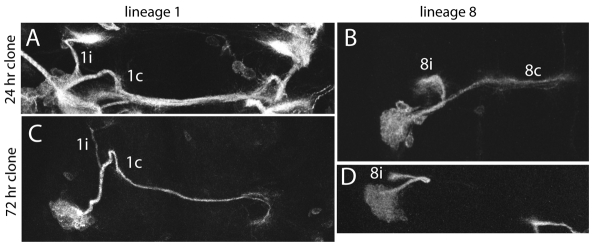
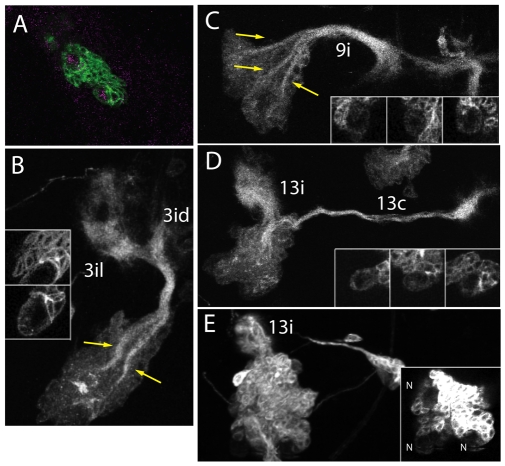
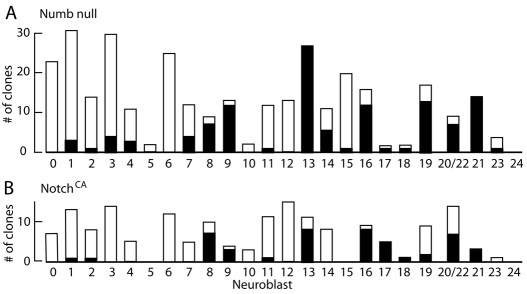
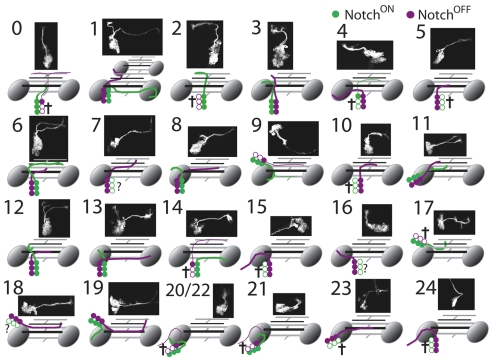
The secondary neurons generated in the thoracic central nervous system of Drosophila arise from a hemisegmental set of 25 neuronal stem cells, the neuroblasts (NBs). Each NB undergoes repeated asymmetric divisions to produce a series of smaller ganglion mother cells (GMCs), which typically divide once to form two daughter neurons. We find that the two daughters of the GMC consistently have distinct fates. Using both loss-of-function and gain-of-function approaches, we examined the role of Notch signaling in establishing neuronal fates within all of the thoracic secondary lineages. In all cases, the 'A' (Notch(ON)) sibling assumes one fate and the 'B' (Notch(OFF)) sibling assumes another, and this relationship holds throughout the neurogenic period, resulting in two major neuronal classes: the A and B hemilineages. Apparent monotypic lineages typically result from the death of one sibling throughout the lineage, resulting in a single, surviving hemilineage. Projection neurons are predominantly from the B hemilineages, whereas local interneurons are typically from A hemilineages. Although sibling fate is dependent on Notch signaling, it is not necessarily dependent on numb, a gene classically involved in biasing Notch activation. When Numb was removed at the start of larval neurogenesis, both A and B hemilineages were still generated, but by the start of the third larval instar, the removal of Numb resulted in all neurons assuming the A fate. The need for Numb to direct Notch signaling correlated with a decrease in NB cell cycle time and may be a means for coping with multiple sibling pairs simultaneously undergoing fate decisions.
Figures
Similar articles
-
On the roles of Notch, Delta, kuzbanian, and inscuteable during the development of Drosophila embryonic neuroblast lineages.Dev Biol. 2009 Dec 15;336(2):156-68. doi: 10.1016/j.ydbio.2009.09.030. Epub 2009 Sep 25. Dev Biol. 2009. PMID: 19782677
-
Inscuteable maintains type I neuroblast lineage identity via Numb/Notch signaling in the Drosophila larval brain.J Genet Genomics. 2017 Mar 20;44(3):151-162. doi: 10.1016/j.jgg.2017.02.005. Epub 2017 Mar 6. J Genet Genomics. 2017. PMID: 28325554
-
Notch regulates the generation of diverse cell types from the lateral lineage of Drosophila antennal lobe.J Neurogenet. 2010 Mar;24(1):42-53. doi: 10.3109/01677060903582202. J Neurogenet. 2010. PMID: 20148759
-
Generating neuronal diversity in the Drosophila central nervous system: a view from the ganglion mother cells.Dev Dyn. 2005 Mar;232(3):609-16. doi: 10.1002/dvdy.20273. Dev Dyn. 2005. PMID: 15704126 Review.
-
Asymmetric segregation of Numb: a mechanism for neural specification from Drosophila to mammals.Nat Neurosci. 2002 Dec;5(12):1265-9. doi: 10.1038/nn1202-1265. Nat Neurosci. 2002. PMID: 12447381 Review.
Cited by
-
Lineage analysis of Drosophila lateral antennal lobe neurons reveals notch-dependent binary temporal fate decisions.PLoS Biol. 2012;10(11):e1001425. doi: 10.1371/journal.pbio.1001425. Epub 2012 Nov 20. PLoS Biol. 2012. PMID: 23185131 Free PMC article.
-
Combinatorial action of Grainyhead, Extradenticle and Notch in regulating Hox mediated apoptosis in Drosophila larval CNS.PLoS Genet. 2017 Oct 12;13(10):e1007043. doi: 10.1371/journal.pgen.1007043. eCollection 2017 Oct. PLoS Genet. 2017. PMID: 29023471 Free PMC article.
-
Genetic and Environmental Control of Neurodevelopmental Robustness in Drosophila.PLoS One. 2016 May 25;11(5):e0155957. doi: 10.1371/journal.pone.0155957. eCollection 2016. PLoS One. 2016. PMID: 27223118 Free PMC article.
-
Transvection is common throughout the Drosophila genome.Genetics. 2012 Aug;191(4):1129-41. doi: 10.1534/genetics.112.140475. Epub 2012 May 29. Genetics. 2012. PMID: 22649078 Free PMC article.
-
Temporal patterning of Drosophila medulla neuroblasts controls neural fates.Nature. 2013 Jun 27;498(7455):456-62. doi: 10.1038/nature12319. Epub 2013 Jun 19. Nature. 2013. PMID: 23783517 Free PMC article.
References
-
- Almeida M. S., Bray S. J. (2005). Regulation of post-embryonic neurogenesis by Drosophila Grainyhead. Mech. Dev. 1212, 1282-1293 - PubMed
-
- Bailey A. M., Posakony J. W. (1995). Suppressor of hairless directly activates transcription of enhancer of split complex genes in response to Notch receptor activity. Genes Dev. 9, 2609-2622 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
