

Dynamics of Delta/Notch signaling on endomesoderm segregation in the sea urchin embryo
- PMID: 20023163
- PMCID: PMC2796929
- DOI: 10.1242/dev.044149
Dynamics of Delta/Notch signaling on endomesoderm segregation in the sea urchin embryo
Abstract
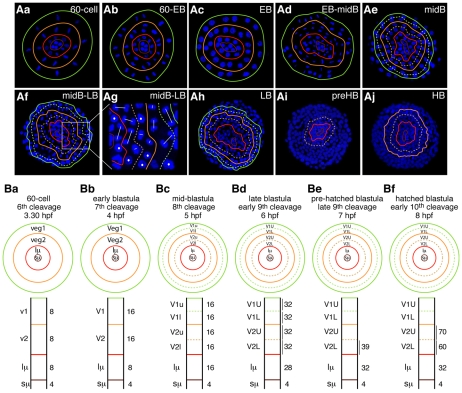
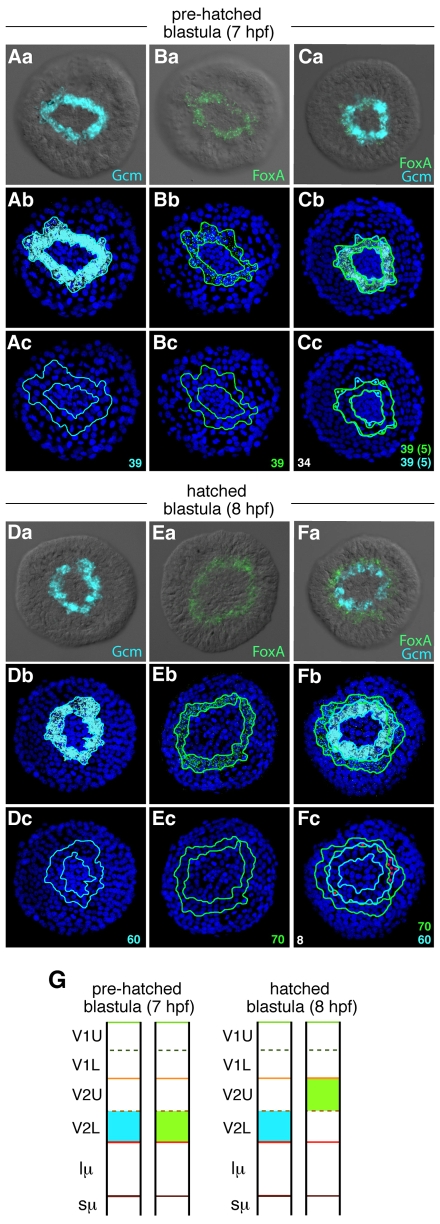
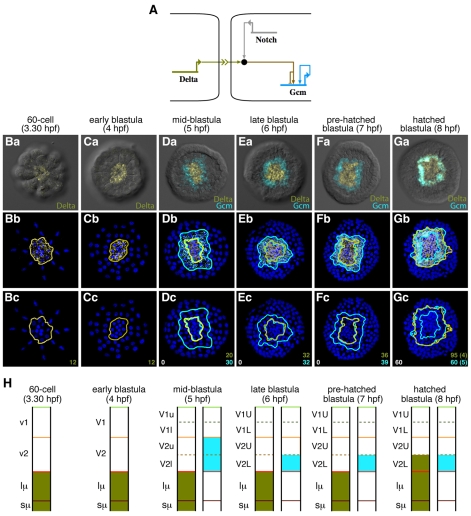
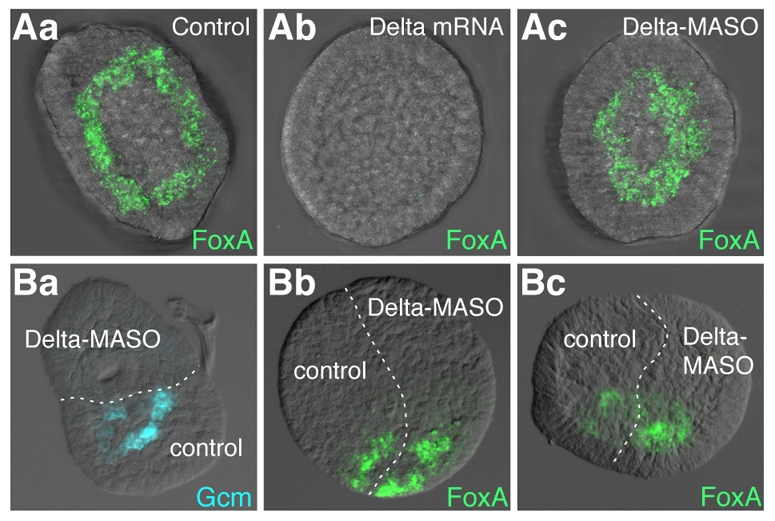
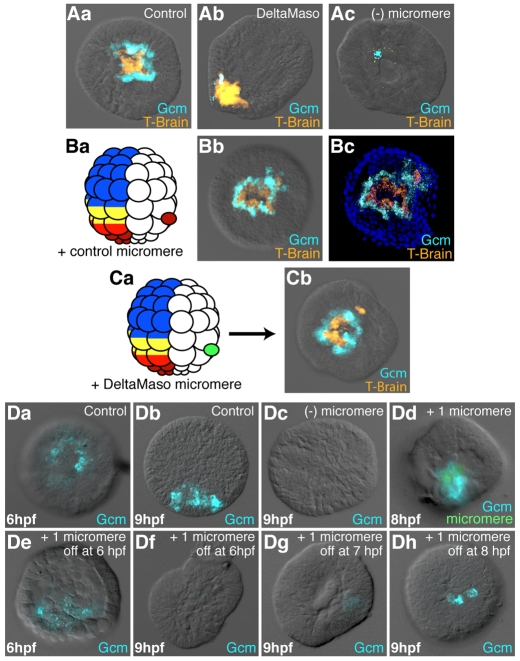
Endomesoderm is the common progenitor of endoderm and mesoderm early in the development of many animals. In the sea urchin embryo, the Delta/Notch pathway is necessary for the diversification of this tissue, as are two early transcription factors, Gcm and FoxA, which are expressed in mesoderm and endoderm, respectively. Here, we provide a detailed lineage analysis of the cleavages leading to endomesoderm segregation, and examine the expression patterns and the regulatory relationships of three known regulators of this cell fate dichotomy in the context of the lineages. We observed that endomesoderm segregation first occurs at hatched blastula stage. Prior to this stage, Gcm and FoxA are co-expressed in the same cells, whereas at hatching these genes are detected in two distinct cell populations. Gcm remains expressed in the most vegetal endomesoderm descendant cells, while FoxA is downregulated in those cells and activated in the above neighboring cells. Initially, Delta is expressed exclusively in the micromeres, where it is necessary for the most vegetal endomesoderm cell descendants to express Gcm and become mesoderm. Our experiments show a requirement for a continuous Delta input for more than two cleavages (or about 2.5 hours) before Gcm expression continues in those cells independently of further Delta input. Thus, this study provides new insights into the timing mechanisms and the molecular dynamics of endomesoderm segregation during sea urchin embryogenesis and into the mode of action of the Delta/Notch pathway in mediating mesoderm fate.
Figures
References
-
- Cameron R. A., Fraser S. E., Britten R. J., Davidson E. H. (1991). Macromere cell fates during sea urchin development. Development 113, 1085-1091 - PubMed
-
- Croce J., Lhomond G., Gache C. (2003). Coquillette, a sea urchin T-box gene of the Tbx2 subfamily, is expressed asymmetrically along the oral-aboral axis of the embryo and is involved in skeletogenesis. Mech. Dev. 120, 561-572 - PubMed
-
- Davidson E. H., Rast J. P., Oliveri P., Ransick A., Calestani C., Yuh C. H., Minokawa T., Amore G., Hinman V., Arenas-Mena C., et al. (2002). A genomic regulatory network for development. Science 295, 1669-1678 - PubMed
-
- Ettensohn C. A., McClay D. R. (1988). Cell lineage conversion in the sea urchin embryo. Dev. Biol. 125, 396-409 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
