

Arabidopsis formin3 directs the formation of actin cables and polarized growth in pollen tubes
- PMID: 20023198
- PMCID: PMC2814512
- DOI: 10.1105/tpc.109.068700
Arabidopsis formin3 directs the formation of actin cables and polarized growth in pollen tubes
Abstract
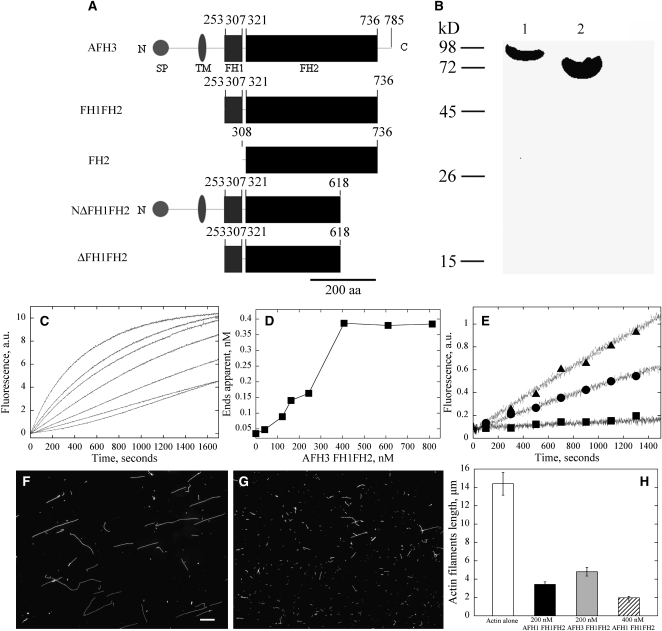
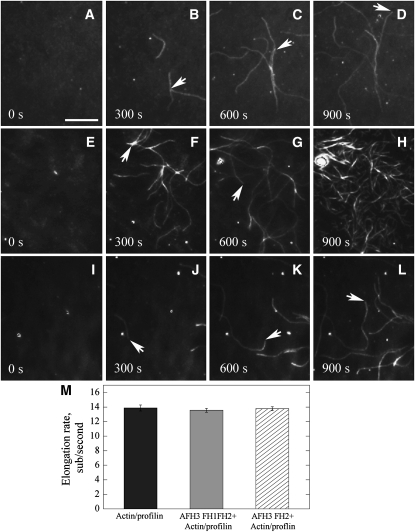
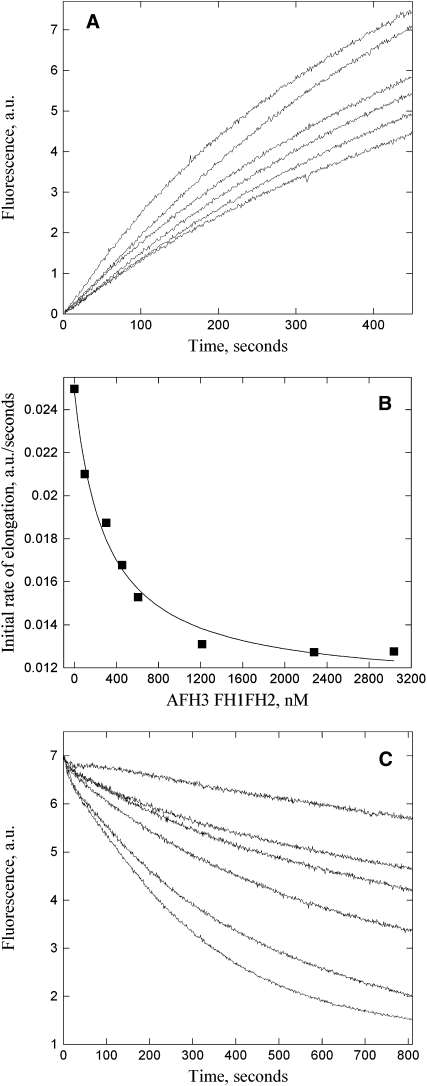
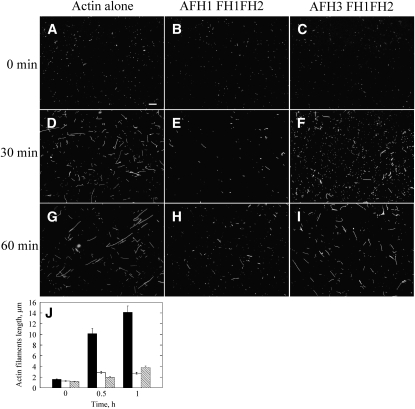
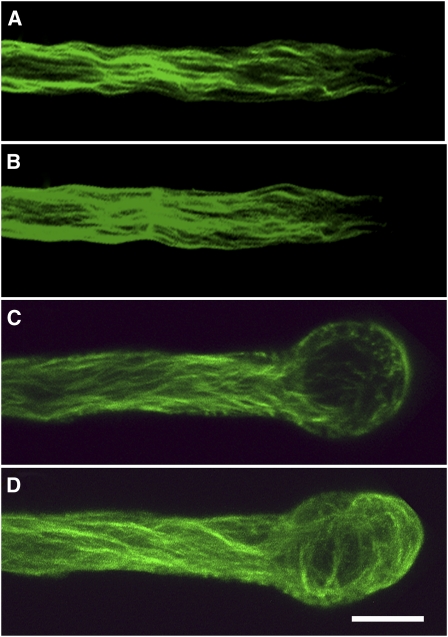
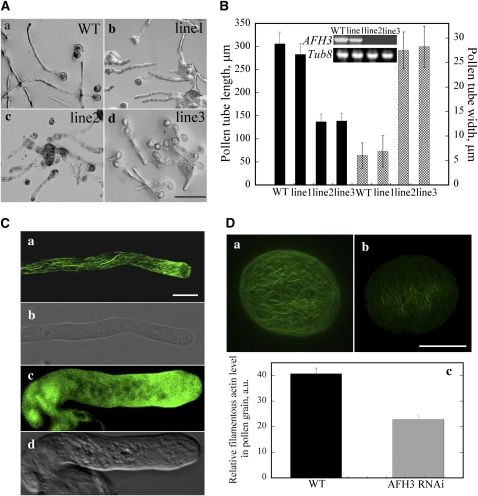
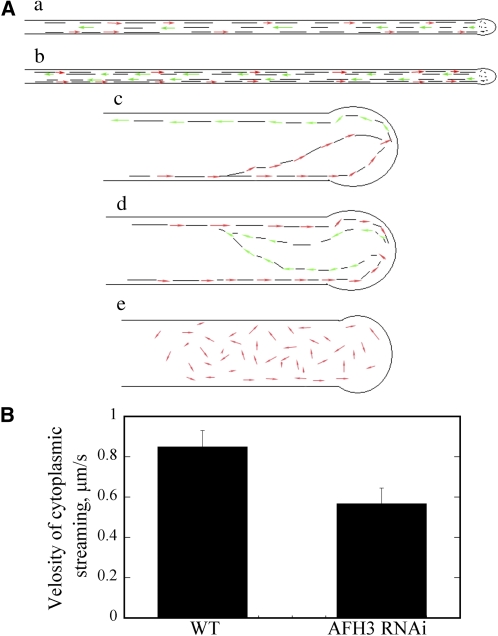
Cytoplasmic actin cables are the most prominent actin structures in plant cells, but the molecular mechanism underlying their formation is unknown. The function of these actin cables, which are proposed to modulate cytoplasmic streaming and intracellular movement of many organelles in plants, has not been studied by genetic means. Here, we show that Arabidopsis thaliana formin3 (AFH3) is an actin nucleation factor responsible for the formation of longitudinal actin cables in pollen tubes. The Arabidopsis AFH3 gene encodes a 785-amino acid polypeptide, which contains a formin homology 1 (FH1) and a FH2 domain. In vitro analysis revealed that the AFH3 FH1FH2 domains interact with the barbed end of actin filaments and have actin nucleation activity in the presence of G-actin or G actin-profilin. Overexpression of AFH3 in tobacco (Nicotiana tabacum) pollen tubes induced excessive actin cables, which extended into the tubes' apices. Specific downregulation of AFH3 eliminated actin cables in Arabidopsis pollen tubes and reduced the level of actin polymers in pollen grains. This led to the disruption of the reverse fountain streaming pattern in pollen tubes, confirming a role for actin cables in the regulation of cytoplasmic streaming. Furthermore, these tubes became wide and short and swelled at their tips, suggesting that actin cables may regulate growth polarity in pollen tubes. Thus, AFH3 regulates the formation of actin cables, which are important for cytoplasmic streaming and polarized growth in pollen tubes.
Figures
Similar articles
-
Arabidopsis actin-depolymerizing factor7 severs actin filaments and regulates actin cable turnover to promote normal pollen tube growth.Plant Cell. 2013 Sep;25(9):3405-23. doi: 10.1105/tpc.113.117820. Epub 2013 Sep 20. Plant Cell. 2013. PMID: 24058157 Free PMC article.
-
Arabidopsis FIMBRIN5, an actin bundling factor, is required for pollen germination and pollen tube growth.Plant Cell. 2010 Nov;22(11):3745-63. doi: 10.1105/tpc.110.080283. Epub 2010 Nov 23. Plant Cell. 2010. PMID: 21098731 Free PMC article.
-
Arabidopsis villins promote actin turnover at pollen tube tips and facilitate the construction of actin collars.Plant Cell. 2013 May;25(5):1803-17. doi: 10.1105/tpc.113.110940. Epub 2013 May 28. Plant Cell. 2013. PMID: 23715472 Free PMC article.
-
The function of actin-binding proteins in pollen tube growth.Protoplasma. 2007;230(3-4):171-82. doi: 10.1007/s00709-006-0231-x. Epub 2007 Apr 24. Protoplasma. 2007. PMID: 17458632 Review.
-
Actin-myosin XI: an intracellular control network in plants.Biochem Biophys Res Commun. 2018 Nov 25;506(2):403-408. doi: 10.1016/j.bbrc.2017.12.169. Epub 2018 Jan 5. Biochem Biophys Res Commun. 2018. PMID: 29307817 Review.
Cited by
-
Two tomato GDP-D-mannose epimerase isoforms involved in ascorbate biosynthesis play specific roles in cell wall biosynthesis and development.J Exp Bot. 2016 Aug;67(15):4767-77. doi: 10.1093/jxb/erw260. Epub 2016 Jul 5. J Exp Bot. 2016. PMID: 27382114 Free PMC article.
-
Class II formin targeting to the cell cortex by binding PI(3,5)P(2) is essential for polarized growth.J Cell Biol. 2012 Jul 23;198(2):235-50. doi: 10.1083/jcb.201112085. Epub 2012 Jul 16. J Cell Biol. 2012. PMID: 22801781 Free PMC article.
-
Arabidopsis FIM5 decorates apical actin filaments and regulates their organization in the pollen tube.J Exp Bot. 2016 May;67(11):3407-17. doi: 10.1093/jxb/erw160. Epub 2016 Apr 25. J Exp Bot. 2016. PMID: 27117336 Free PMC article.
-
OsFH3 Encodes a Type II Formin Required for Rice Morphogenesis.Int J Mol Sci. 2021 Dec 9;22(24):13250. doi: 10.3390/ijms222413250. Int J Mol Sci. 2021. PMID: 34948047 Free PMC article.
-
Phospholipids: molecules regulating cytoskeletal organization in plant abiotic stress tolerance.Plant Signal Behav. 2014;9(3):e28337. doi: 10.4161/psb.28337. Epub 2014 Jan 1. Plant Signal Behav. 2014. PMID: 24589893 Free PMC article. Review.
References
-
- Banno, H., and Chua, N.H. (2000). Characterization of the arabidopsis formin-like protein AFH1 and its interacting protein. Plant Cell Physiol. 41 617–626. - PubMed
-
- Bechtold, N., and Pelletier, G. (1998). In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 82 259–266. - PubMed
-
- Blanchoin, L., Amann, K.J., Higgs, H.N., Marchand, J.B., Kaiser, D.A., and Pollard, T.D. (2000). Direct observation of dendritic actin filament networks nucleated by Arp2/3 complex and WASP/Scar proteins. Nature 404 1007–1011. - PubMed
-
- Blanchoin, L., and Staiger, C.J. (2008). Plant formins: Diverse isoforms and unique molecular mechanism. Biochim. Biophys. Acta doi:10.1016/j.bbamcr.2008.09.015. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
