

Distinct roles for LFA-1 affinity regulation during T-cell adhesion, diapedesis, and interstitial migration in lymph nodes
- PMID: 20023213
- PMCID: PMC2830762
- DOI: 10.1182/blood-2009-08-237917
Distinct roles for LFA-1 affinity regulation during T-cell adhesion, diapedesis, and interstitial migration in lymph nodes
Abstract
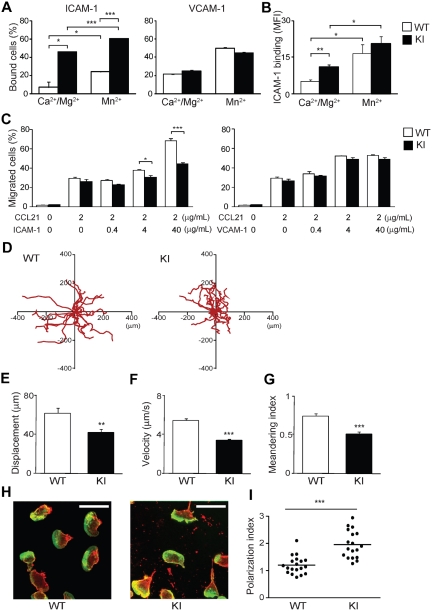
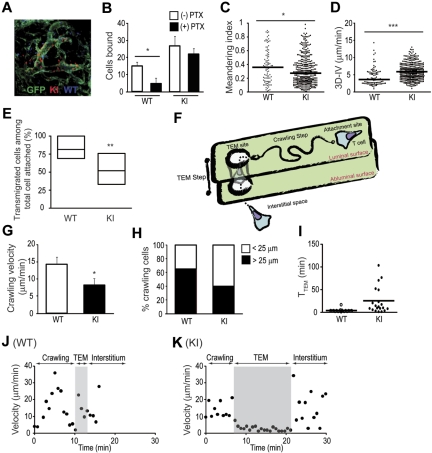
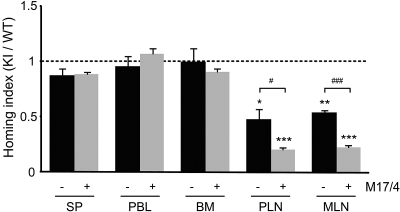
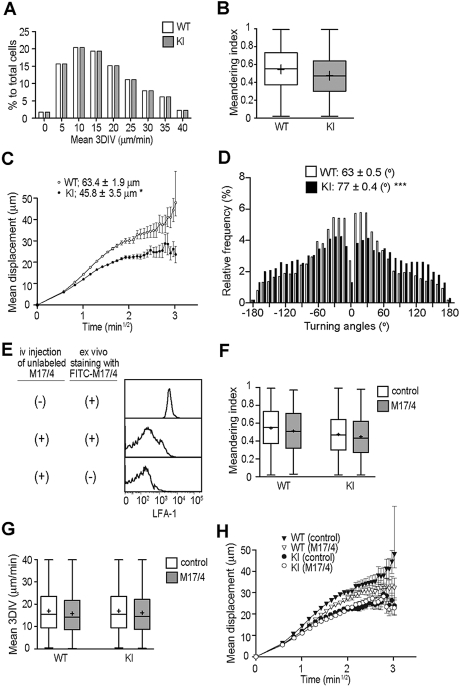
During the course of homing to lymph nodes (LNs), T cells undergo a multistep adhesion cascade that culminates in a lymphocyte function-associated antigen 1 (LFA-1)-dependent firm adhesion to the luminal surface of high endothelial venules (HEVs). The importance of LFA-1 affinity regulation in supporting T-cell arrest on HEVs has been well established, however, its importance in the postadhesion phase, which involves intraluminal crawling and diapedesis to the extravascular space, remains elusive. Here we have shown that LFA-1 affinity needs to be appropriately regulated to support these essential steps in the homing cascade. Genetically engineered T cells that were unable to properly down-regulate LFA-1 affinity underwent enhanced, chemokine-independent arrest in HEVs but showed perturbed intravascular crawling to transmigration sites and compromised diapedesis across HEVs. By contrast, the extravascular migration of T cells was insensitive to the affinity-enhancing LFA-1 mutation. These results highlight the requirement for balanced LFA-1 affinity regulation in intravascular and transvascular, but not extravascular, T-cell migration in LNs.
Figures



References
-
- von Andrian UH, Mackay CR. T-cell function and migration: two sides of the same coin. N Engl J Med. 2000;343(14):1020–1034. - PubMed
-
- Mempel TR, Scimone ML, Mora JR, von Andrian UH. In vivo imaging of leukocyte trafficking in blood vessels and tissues. Curr Opin Immunol. 2004;16(4):406–417. - PubMed
-
- Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7(9):678–689. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI061663/AI/NIAID NIH HHS/United States
- AR42689/AR/NIAMS NIH HHS/United States
- HL56949/HL/NHLBI NIH HHS/United States
- AI078897/AI/NIAID NIH HHS/United States
- R01 AI063421/AI/NIAID NIH HHS/United States
- AI063421/AI/NIAID NIH HHS/United States
- P30 AR042689/AR/NIAMS NIH HHS/United States
- P01 AI078897/AI/NIAID NIH HHS/United States
- AI069259/AI/NIAID NIH HHS/United States
- P01 HL048675/HL/NHLBI NIH HHS/United States
- P01 HL056949/HL/NHLBI NIH HHS/United States
- AI072252/AI/NIAID NIH HHS/United States
- R01 AI061663/AI/NIAID NIH HHS/United States
- R01 AI072252/AI/NIAID NIH HHS/United States
- R01 AI069259/AI/NIAID NIH HHS/United States
- HL048675/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
