

Pulling force generated by interacting SNAREs facilitates membrane hemifusion
- PMID: 20023730
- PMCID: PMC2865556
- DOI: 10.1039/b900685k
Pulling force generated by interacting SNAREs facilitates membrane hemifusion
Abstract
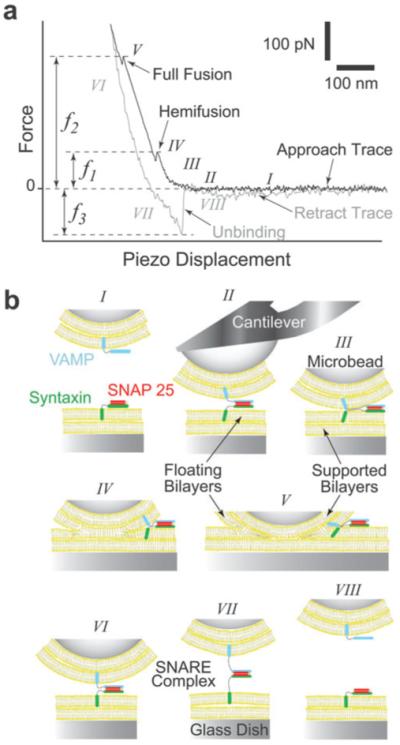
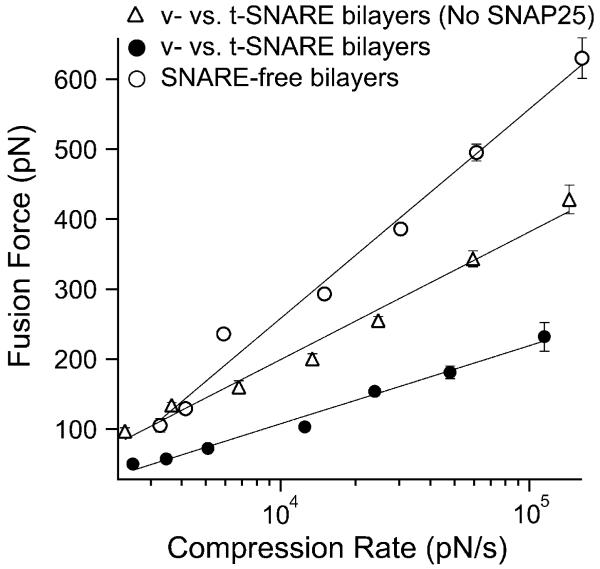
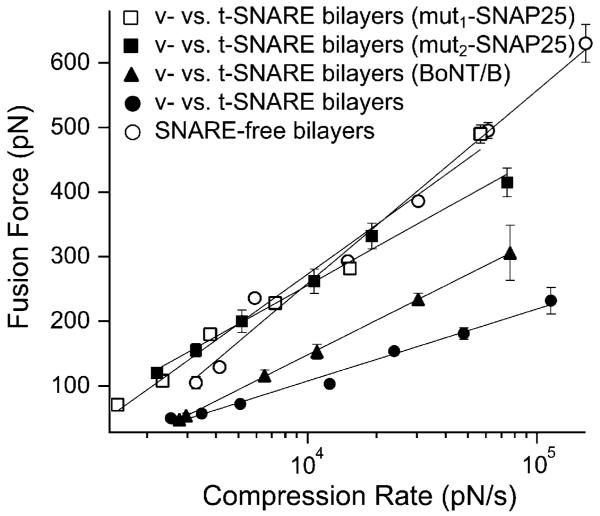
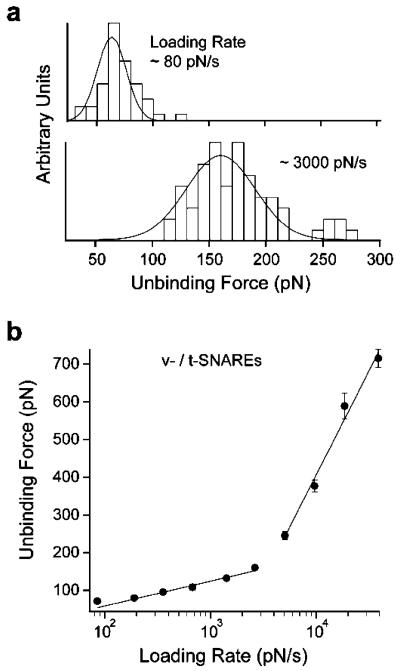
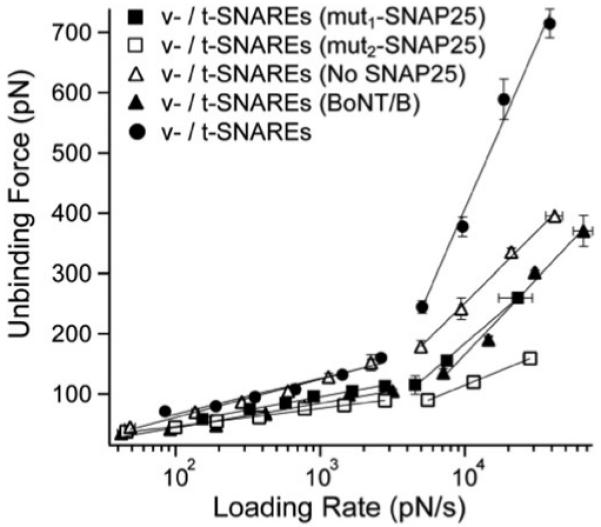
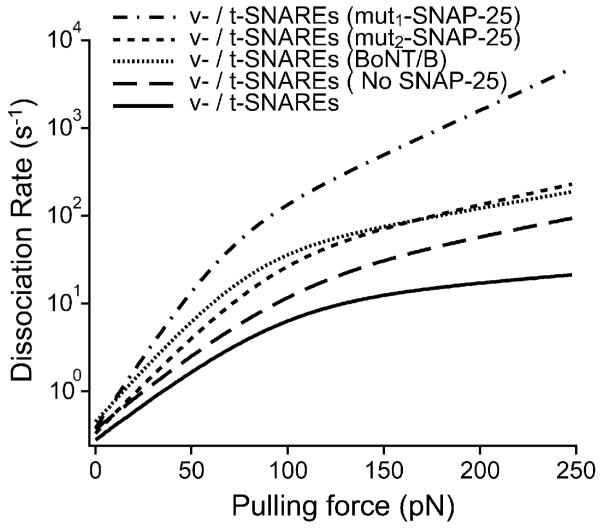
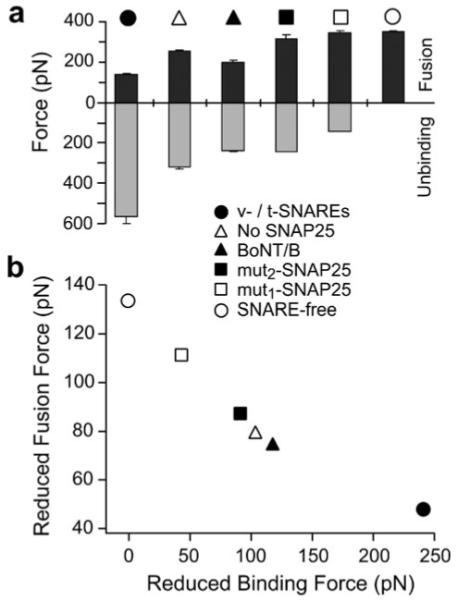
In biological systems, membrane fusion is mediated by specialized proteins. Although soluble N-ethylmaleimide-sensitive factor attachment protein (SNAP) receptors (SNAREs) provide the minimal molecular machinery required to drive membrane fusion, the precise mechanism for SNARE-mediated fusion remains to be established. Here, we used atomic force microscope (AFM) spectroscopy to determine whether the pulling force generated by interacting SNAREs is directly coupled to membrane fusion. The mechanical strength of the SNARE binding interaction was determined by single molecule force measurements. It was revealed that the forced unbinding of the SNARE complex formed between opposing (trans) bilayers involves two activation barriers; where the steep inner barrier governs the transition from the bound to an intermediate state and the outer barrier governs the transition between the intermediate and the unbound state. Moreover, truncation of either SNAP-25 or VAMP 2 reduced the slope of the inner barrier significantly and, consequently, reduced the pulling strength of the SNARE complex; thus, suggesting that the inner barrier determines the binding strength of the SNARE complex. In parallel, AFM compression force measurements revealed that truncated SNAREs were less efficient than native SNAREs in facilitating hemifusion of the apposed bilayers. Together, these findings reveal a mechanism by which a pulling force generated by interacting trans-SNAREs reduces the slope of the hemifusion barrier and, subsequently, facilitates hemifusion and makes the membranes more prone to fusion.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
