

Testosterone metabolites differentially maintain adult morphology in a sexually dimorphic neuromuscular system
- PMID: 20024940
- PMCID: PMC2905164
- DOI: 10.1002/dneu.20780
Testosterone metabolites differentially maintain adult morphology in a sexually dimorphic neuromuscular system
Abstract
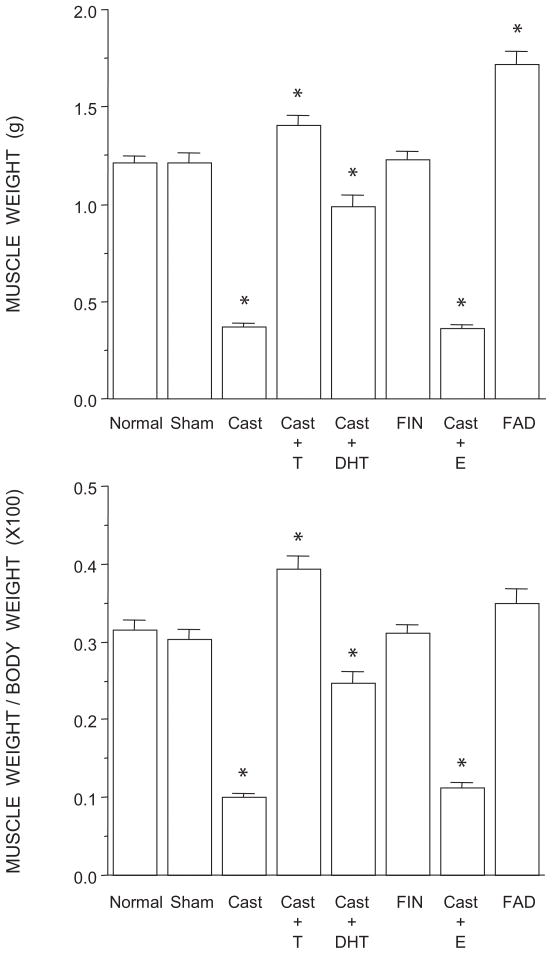
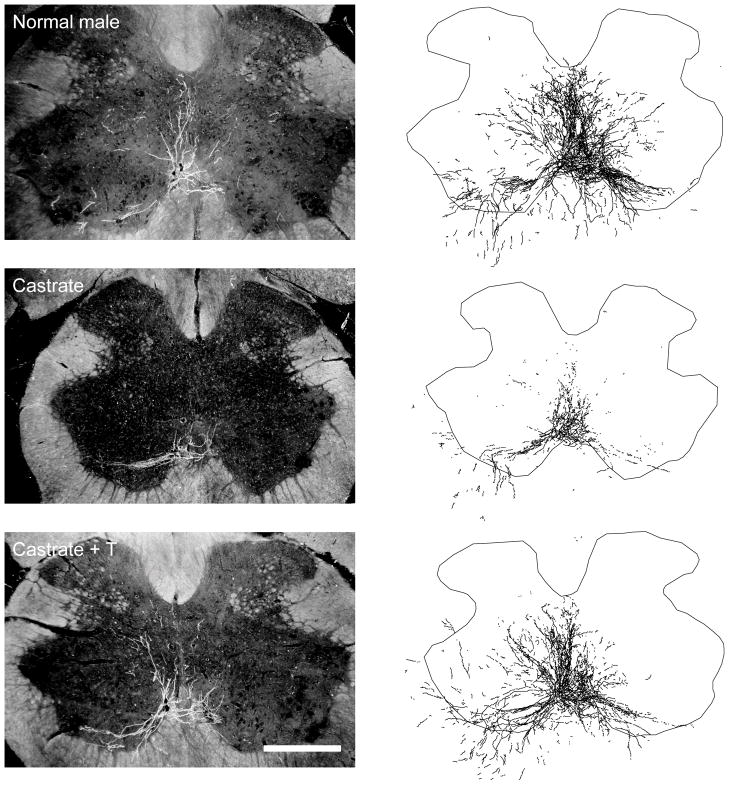
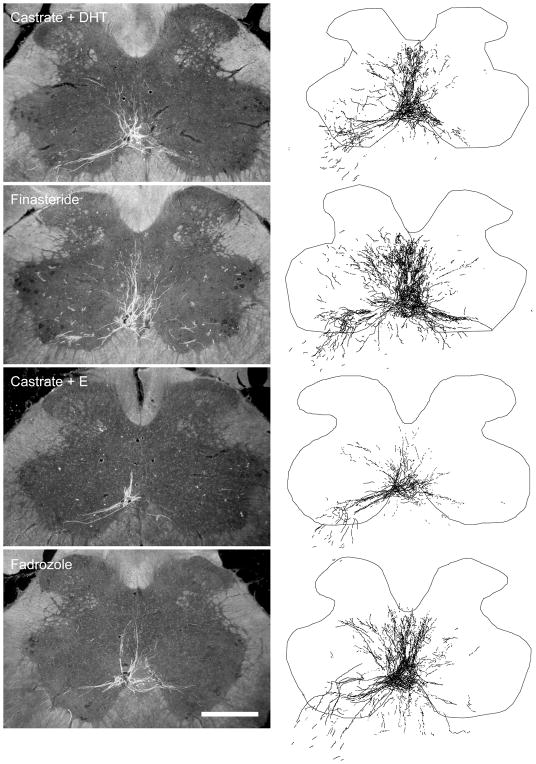
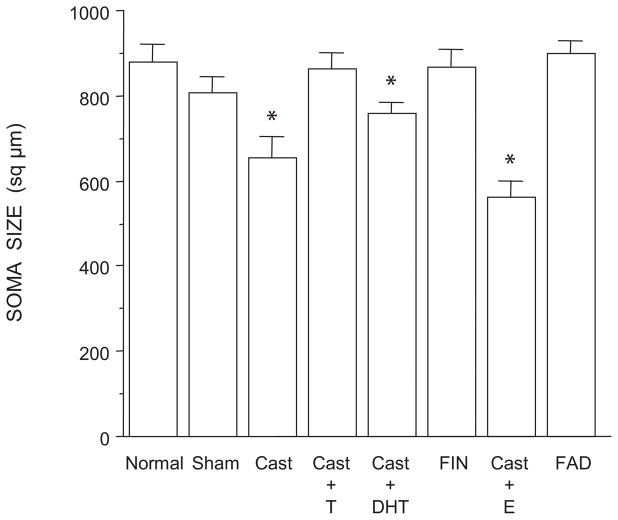
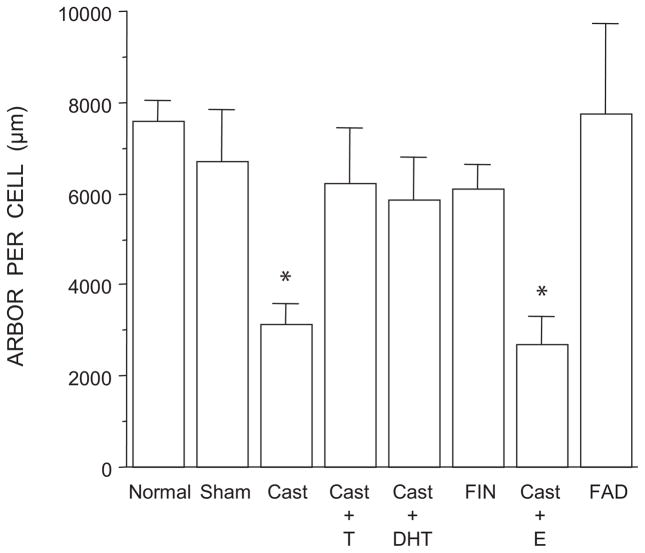
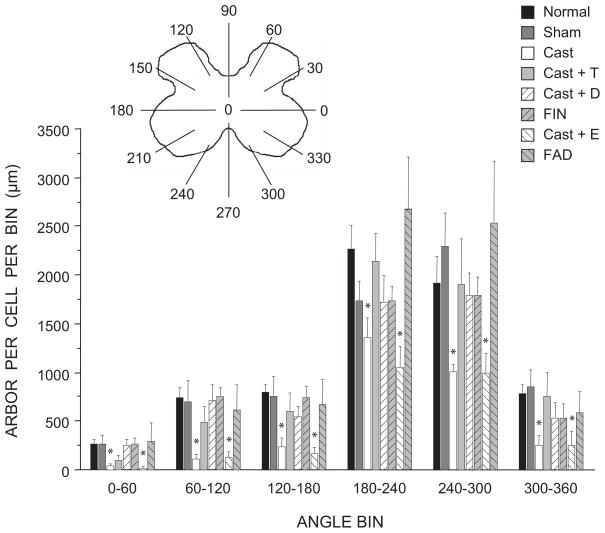
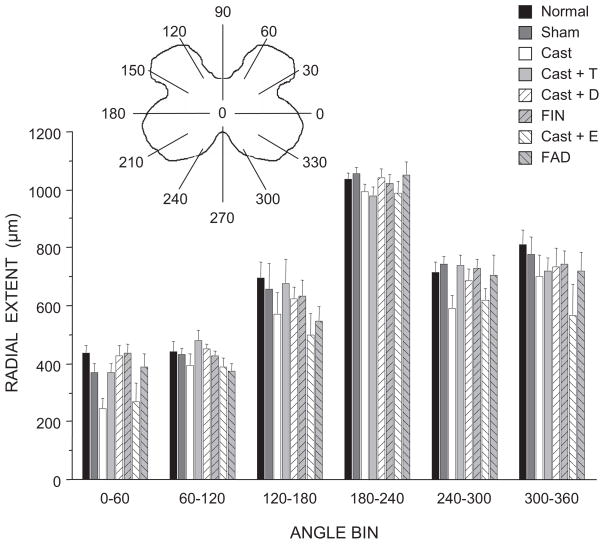
The lumbar spinal cord of rats contains the sexually dimorphic, steroid-sensitive spinal nucleus of the bulbocavernosus (SNB). Androgens are necessary for the development of the SNB neuromuscular system, and in adulthood, continue to influence the morphology and function of the motoneurons and their target musculature. However, estrogens are also involved in the development of the SNB system, and are capable of maintaining function in adulthood. In this experiment, we assessed the ability of testosterone metabolites, estrogens and nonaromatizable androgens, to maintain neuromuscular morphology in adulthood. Motoneuron and muscle morphology was assessed in adult normal males, sham-castrated males, castrated males treated with testosterone, dihydrotestosterone, estradiol, or left untreated, and gonadally intact males treated with the 5alpha-reductase inhibitor finasteride or the aromatase inhibitor fadrozole. After 6 weeks of treatment, SNB motoneurons were retrogradely labeled with cholera toxin-HRP and reconstructed in three dimensions. Castration resulted in reductions in SNB target muscle size, soma size, and dendritic morphology. Testosterone treatment after castration maintained SNB soma size, dendritic morphology, and elevated target muscle size; dihydrotestosterone treatment also maintained SNB dendritic length, but was less effective than testosterone in maintaining both SNB soma size and target muscle weight. Treatment of intact males with finasteride or fadrozole did not alter the morphology of SNB motoneurons or their target muscles. In contrast, estradiol treatment was completely ineffective in preventing castration-induced atrophy of the SNB neuromuscular system. Together, these results suggest that the maintenance of adult motoneuron or muscle morphology is strictly mediated by androgens.
Figures
Similar articles
-
Estrogenic support of motoneuron dendritic growth via the neuromuscular periphery in a sexually dimorphic motor system.J Neurobiol. 2006 Aug;66(9):962-76. doi: 10.1002/neu.20274. J Neurobiol. 2006. PMID: 16779828
-
Aromatase inhibition reduces dendritic growth in a sexually dimorphic rat spinal nucleus.J Neurobiol. 1999 Feb 15;38(3):301-12. J Neurobiol. 1999. PMID: 10022574
-
Trophic effects of brain-derived neurotrophic factor blockade in an androgen-sensitive neuromuscular system.Endocrinology. 2010 Nov;151(11):5337-48. doi: 10.1210/en.2010-0799. Epub 2010 Sep 22. Endocrinology. 2010. PMID: 20861229 Free PMC article.
-
The spinal nucleus of the bulbocavernosus: firsts in androgen-dependent neural sex differences.Horm Behav. 2008 May;53(5):596-612. doi: 10.1016/j.yhbeh.2007.11.008. Epub 2007 Nov 28. Horm Behav. 2008. PMID: 18191128 Free PMC article. Review.
-
Cellular analyses of hormone influence on motoneuronal development and function.J Neurobiol. 1986 May;17(3):157-76. doi: 10.1002/neu.480170304. J Neurobiol. 1986. PMID: 3519862 Review.
Cited by
-
Androgen action at the target musculature regulates brain-derived neurotrophic factor protein in the spinal nucleus of the bulbocavernosus.Dev Neurobiol. 2013 Aug;73(8):587-98. doi: 10.1002/dneu.22083. Epub 2013 Jun 24. Dev Neurobiol. 2013. PMID: 23512738 Free PMC article.
-
Intracrine and myotrophic roles of 5α-reductase and androgens: a review.Med Sci Sports Exerc. 2012 May;44(5):818-26. doi: 10.1249/MSS.0b013e31823bfcbf. Med Sci Sports Exerc. 2012. PMID: 21988936 Free PMC article.
-
Critical period for estrogen-dependent motoneuron dendrite growth is coincident with ERα expression in target musculature.Dev Neurobiol. 2013 Jan;73(1):72-84. doi: 10.1002/dneu.22040. Epub 2012 Jul 27. Dev Neurobiol. 2013. PMID: 22678724 Free PMC article.
-
Castration-induced upregulation of muscle ERα supports estrogen sensitivity of motoneuron dendrites in a sexually dimorphic neuromuscular system.Dev Neurobiol. 2013 Dec;73(12):921-35. doi: 10.1002/dneu.22118. Epub 2013 Oct 7. Dev Neurobiol. 2013. PMID: 23939785 Free PMC article.
-
Protective Effects of Estradiol and Dihydrotestosterone following Spinal Cord Injury.J Neurotrauma. 2018 Mar 15;35(6):825-841. doi: 10.1089/neu.2017.5329. Epub 2018 Jan 11. J Neurotrauma. 2018. PMID: 29132243 Free PMC article.
References
-
- Argente J, Chowen-Breed JA, Steiner RA, Clifton DK. Somatostatin messenger-rna in hypothalamic neurons is increased by testosterone through activation of androgen receptors and not by aromatization to estradiol. Neuroendocrinol. 1990;52:342–349. - PubMed
-
- Bartsch W, Knabbe C, Voigt KD. Regulation and compartmentalization of androgens in rat prostate and muscle. J Steroid Biochem. 1983;19:929–937. - PubMed
-
- Bleisch WV, Harrelson A. Androgens modulate endplate size and Ach receptor density at synapses in rat levator ani muscle. J Neurobiol. 1989;20:189–202. - PubMed
-
- Bleisch WV, Harrelson AL, Luine VN. Testosterone increases acetylcholine receptor number in the “levator ani” muscle of the rat. J Neurobiol. 1982;13:153–161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
