

Two CDC42 paralogues modulate Cryptococcus neoformans thermotolerance and morphogenesis under host physiological conditions
- PMID: 20025659
- PMCID: PMC3685590
- DOI: 10.1111/j.1365-2958.2009.07019.x
Two CDC42 paralogues modulate Cryptococcus neoformans thermotolerance and morphogenesis under host physiological conditions
Abstract
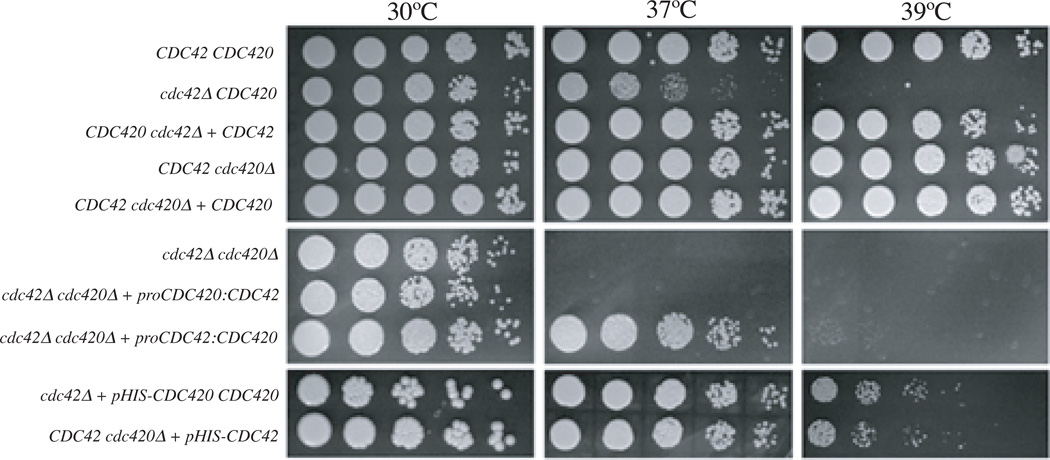
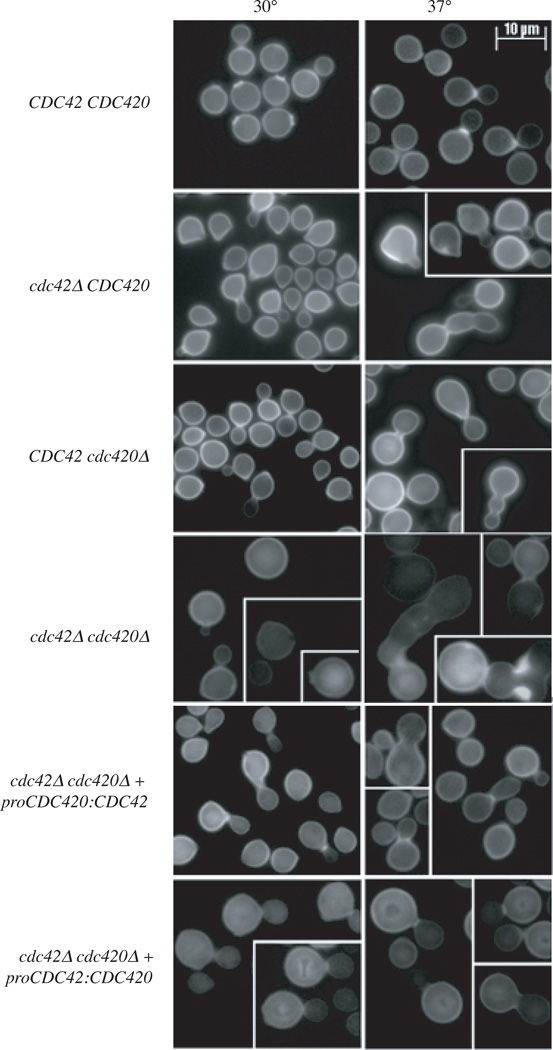
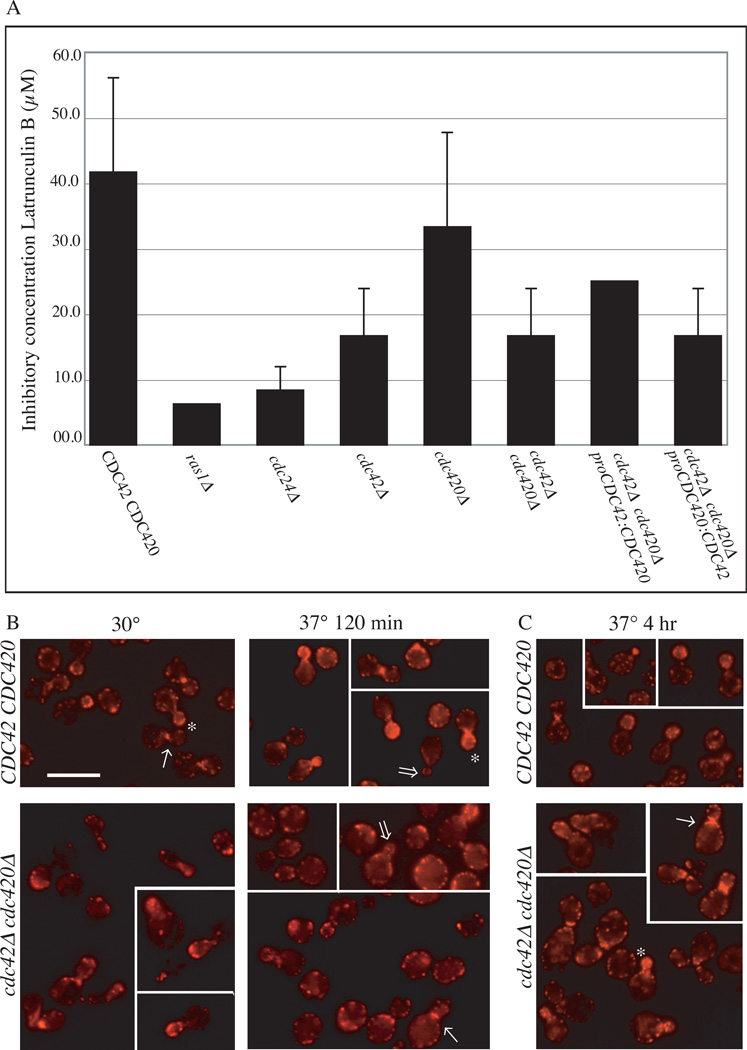
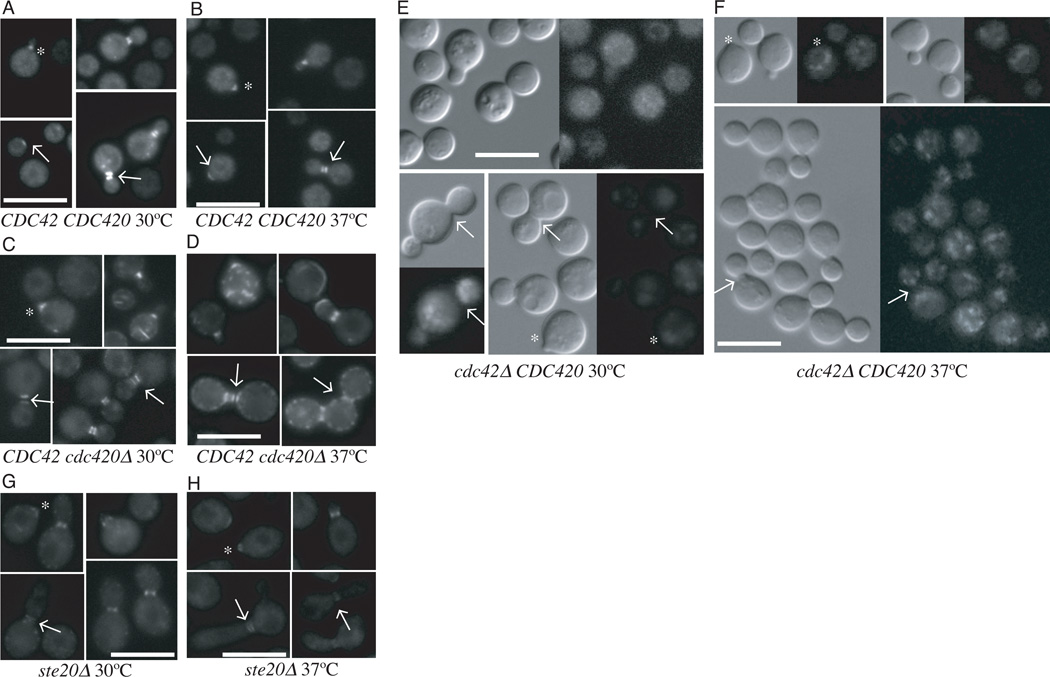
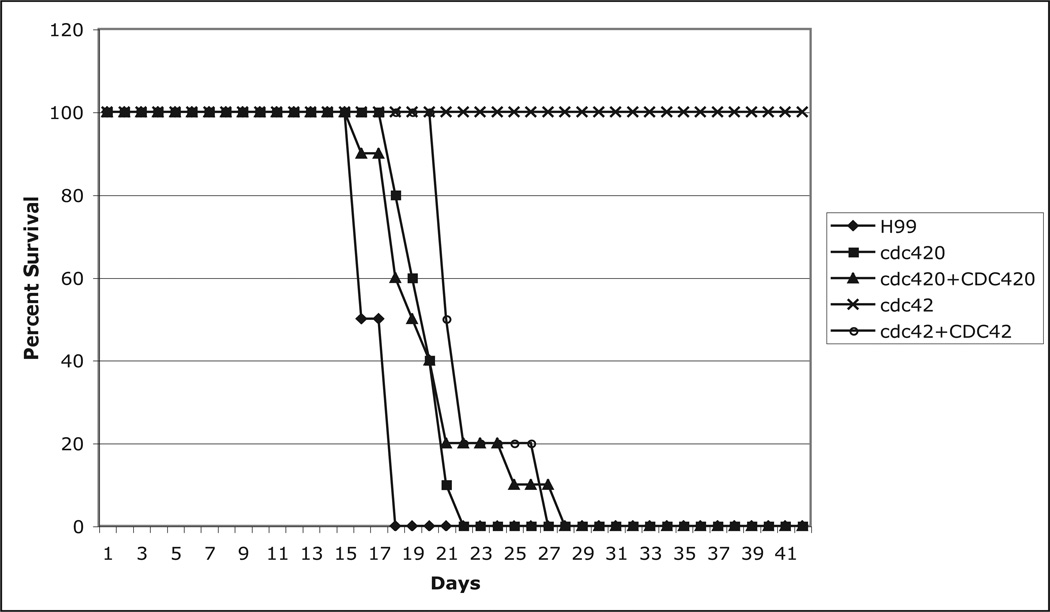
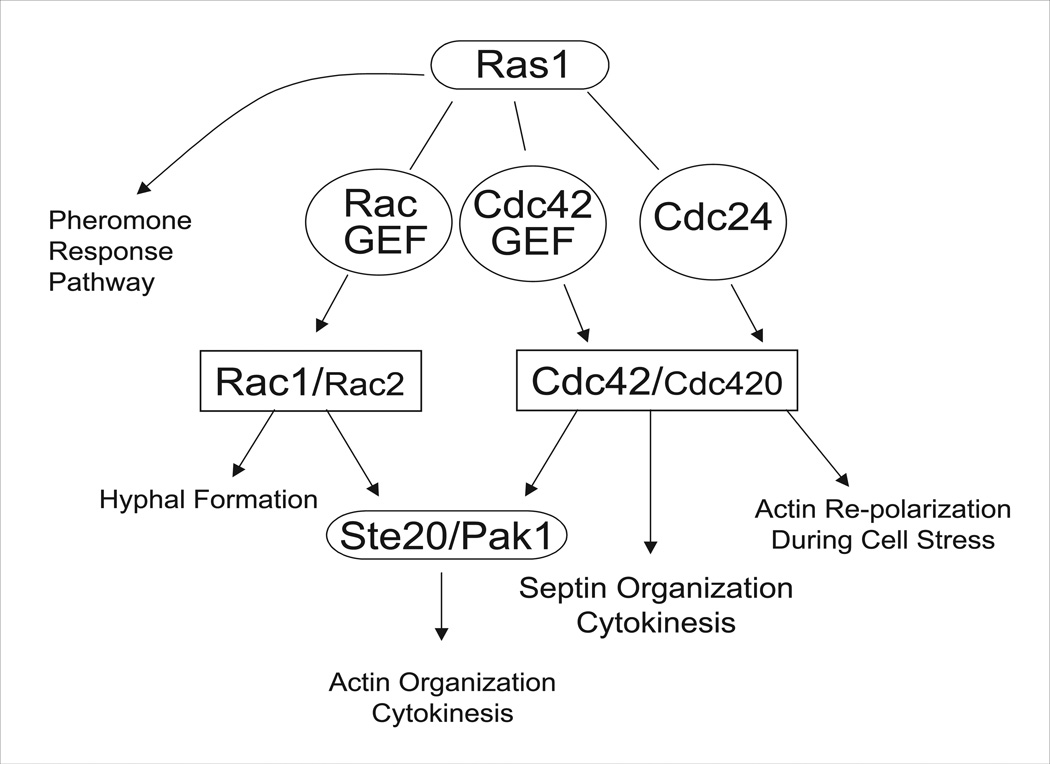
The precise regulation of morphogenesis is a key mechanism by which cells respond to a variety of stresses, including those encountered by microbial pathogens in the host. The polarity protein Cdc42 regulates cellular morphogenesis throughout eukaryotes, and we explore the role of Cdc42 proteins in the host survival of the human fungal pathogen Cryptococcus neoformans. Uniquely, C. neoformans has two functional Cdc42 paralogues, Cdc42 and Cdc420. Here we investigate the contribution of each paralogue to resistance to host stress. In contrast to non-pathogenic model organisms, C. neoformans Cdc42 proteins are not required for viability under non-stress conditions but are required for resistance to high temperature. The paralogues play differential roles in actin and septin organization and act downstream of C. neoformans Ras1 to regulate its morphogenesis sub-pathway, but not its effects on mating. Cdc42, and not Cdc420, is upregulated in response to temperature stress and is required for virulence in a murine model of cryptococcosis. The C. neoformans Cdc42 proteins likely perform complementary functions with other Rho-like GTPases to control cell polarity, septin organization and hyphal transitions that allow survival in the environment and in the host.
Figures



Similar articles
-
Ras1 acts through duplicated Cdc42 and Rac proteins to regulate morphogenesis and pathogenesis in the human fungal pathogen Cryptococcus neoformans.PLoS Genet. 2013;9(8):e1003687. doi: 10.1371/journal.pgen.1003687. Epub 2013 Aug 8. PLoS Genet. 2013. PMID: 23950731 Free PMC article.
-
Two Rac paralogs regulate polarized growth in the human fungal pathogen Cryptococcus neoformans.Fungal Genet Biol. 2013 Aug;57:58-75. doi: 10.1016/j.fgb.2013.05.006. Epub 2013 Jun 5. Fungal Genet Biol. 2013. PMID: 23748012 Free PMC article.
-
A Ras1-Cdc24 signal transduction pathway mediates thermotolerance in the fungal pathogen Cryptococcus neoformans.Mol Microbiol. 2007 Feb;63(4):1118-30. doi: 10.1111/j.1365-2958.2006.05566.x. Mol Microbiol. 2007. PMID: 17233829
-
Cryptococcus neoformans: morphogenesis, infection, and evolution.Infect Genet Evol. 2009 Jul;9(4):401-16. doi: 10.1016/j.meegid.2009.01.013. Epub 2009 Feb 12. Infect Genet Evol. 2009. PMID: 19460306 Review.
-
Cryptococcal Titan Cells: When Yeast Cells Are All Grown up.Curr Top Microbiol Immunol. 2019;422:101-120. doi: 10.1007/82_2018_145. Curr Top Microbiol Immunol. 2019. PMID: 30406867 Review.
Cited by
-
A Fungus-Specific Protein Domain Is Essential for RasA-Mediated Morphogenetic Signaling in Aspergillus fumigatus.mSphere. 2016 Nov 30;1(6):e00234-16. doi: 10.1128/mSphere.00234-16. eCollection 2016 Nov-Dec. mSphere. 2016. PMID: 27921081 Free PMC article.
-
Cryptococcal titan cell formation is regulated by G-protein signaling in response to multiple stimuli.Eukaryot Cell. 2011 Oct;10(10):1306-16. doi: 10.1128/EC.05179-11. Epub 2011 Aug 5. Eukaryot Cell. 2011. PMID: 21821718 Free PMC article.
-
Identification of the Antifungal Metabolite Chaetoglobosin P From Discosia rubi Using a Cryptococcus neoformans Inhibition Assay: Insights Into Mode of Action and Biosynthesis.Front Microbiol. 2020 Jul 28;11:1766. doi: 10.3389/fmicb.2020.01766. eCollection 2020. Front Microbiol. 2020. PMID: 32849391 Free PMC article.
-
Cryptococcus neoformans requires the TVF1 gene for thermotolerance and virulence.Med Mycol. 2023 Oct 5;61(10):myad101. doi: 10.1093/mmy/myad101. Med Mycol. 2023. PMID: 37818721 Free PMC article.
-
Genome-Wide Identification of circRNAs in Pathogenic Basidiomycetous Yeast Cryptococcus neoformans Suggests Conserved circRNA Host Genes over Kingdoms.Genes (Basel). 2018 Feb 26;9(3):118. doi: 10.3390/genes9030118. Genes (Basel). 2018. PMID: 29495353 Free PMC article.
References
-
- Alspaugh JA, Cavallo LM, Perfect JR, Heitman J. RAS1 regulates filamentation, mating and growth at high temperature of Cryptococcus neoformans . Mol Microbiol. 2000;36:352–365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
