

Review
doi: 10.1016/j.molbiopara.2009.12.004.
Epub 2009 Dec 23.
Phospholipid and sphingolipid metabolism in Leishmania
Affiliations
- PMID: 20026359
- PMCID: PMC2815228
- DOI: 10.1016/j.molbiopara.2009.12.004
Item in Clipboard
Review
Phospholipid and sphingolipid metabolism in Leishmania
Mol Biochem Parasitol.
2010 Apr.
Abstract
In many eukaryotes, phospholipids (PLs) and sphingolipids (SLs) are abundant membrane components and reservoirs for important signaling molecules. In Leishmania, the composition, metabolism, and function of PLs and SLs differ significantly from those in mammalian cells. Although only a handful of enzymes have been experimentally characterized, available data suggest many steps of PL/SL metabolism are critical for Leishmania viability and/or virulence, and could be a source for new drug targets. Further studies of genes involved in the synthesis (de novo and salvage) and degradation of PLs and SLs will reveal their diverse effects on Leishmania pathogenesis.
Figures
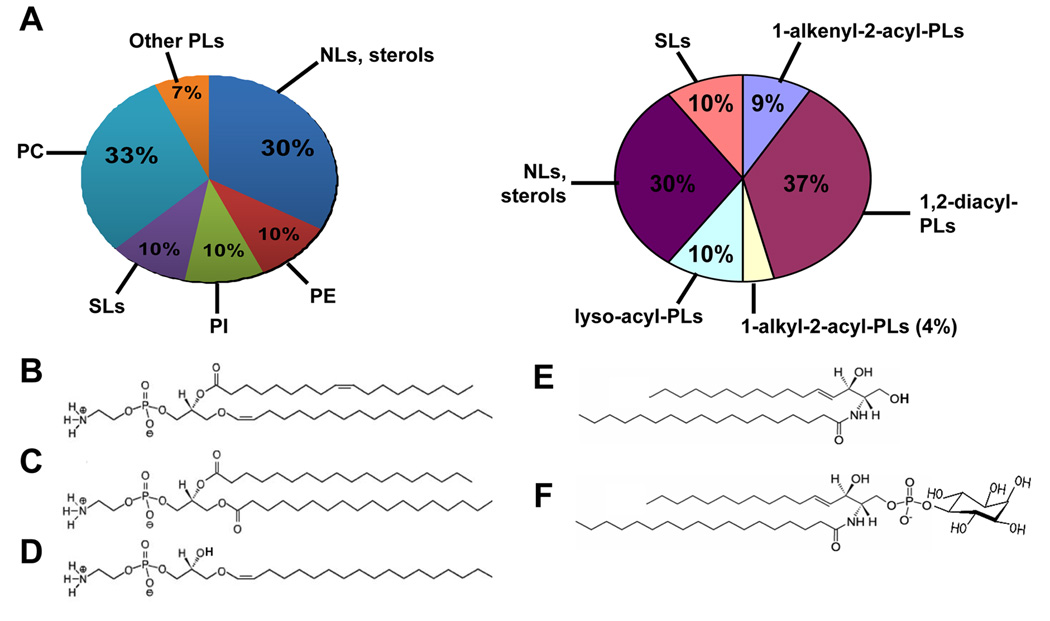
PLs and SLs in L. major. (A) Lipid composition in L. major promastigotes based on the headgroups (left) or anchors (right) of PLs. Structures of each class of lipids were determined by ESI/MS/MS and abundance was estimated as previously described [22, 23, 26]. Other PLs include phosphatidylglycerol (PG), cardiolipin (CL), and phosphatidic acid. (B)–(F) Molecular species of PE and SLs in L. major. (B) A PLE: 1-O-octadec-1’-enyl 2-octadecenoyl sn-glycero-3-phosphoethanolamine (p18:0/18:1-PE). (C) A diacyl PE: 1,2-distearoyl sn-glycero-3-phosphoethanolamine (18:0/18:0-PE). (D) A lyso-PE: 1-O-octadec-1’-enyl 2-lyso sn-glycero-3-phosphoethanolamine (p18:0/lyso-PE). (E) A ceramide: N-stearoylhexadecesphing-4-enine (d16:1/18:0-Cer). (F) An IPC: phosphoryl inositol N-stearoylhexadecesphing-4-enine (d16:1/18:0-IPC).
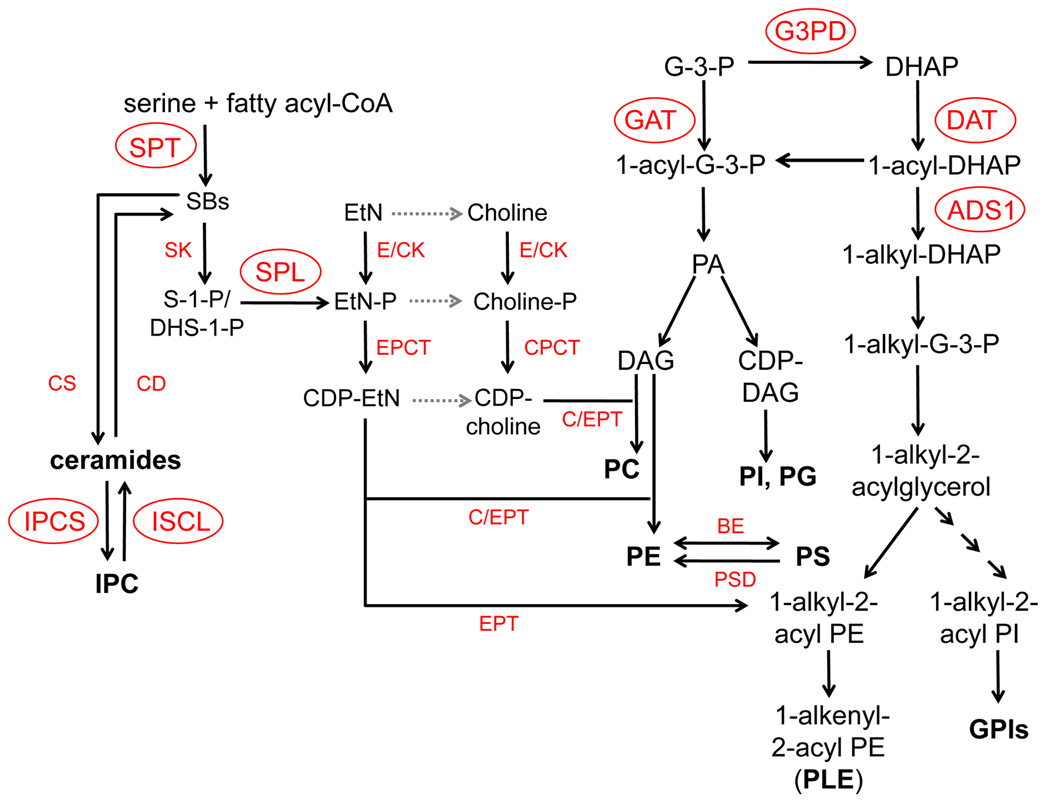
Predicted metabolism of SLs and PLs in Leishmania. DAG: diacylglycerol; G-3-P: glycerol-3-phosphate; DHAP: dihydroxyacetonephosphate. Enzymes of which candidate genes have been identified in the L. major geneDB are indicated. Gray dotted arrows represent pathways where the putative enzymes have yet to be identified. Experimentally characterized enzymes/genes (circled) include DAT (dihydroxyacetone phosphate acyltransferase, system ID: LmjF34.1090), GAT (glycerol-3-phosphate acyltransferase, LmjF03.0080), ADS1 (1-alkyl dihydroxyacetone phosphate synthase 1, LmjF30.0120), G3PD (glycerol-3-phosphate dehydrogenase, LmjF10.0510), SPT (serine palmitoyltransferase, LmjF34.3740 and LmjF35.0320), SPL (sphingosine-1-phosphate lyase, LmjF30.2350), IPCS (IPC synthase, LmjF35.4990), and ISCL (inositol sphingolipidphospholipase C-like, LmjF08.0200). Other enzymes (that have yet to be characterized although their putative genes have been identified from L. major genome) include CS (ceramide synthase); CD (ceramidase); SK (sphingosine kinase); E/CK (ethanolamine/choline kinase); EPCT (ethanolamine-phosphate cytidylyltransferase); CPCT (cholinephosphate cytidylyltransferase); EPT (ethanolamine-specific phosphotransferase); C/EPT (choline/ethanolamine phosphotransferase); BE: (base exchange enzyme or PS synthase); and PSD (PS decarboxylase).
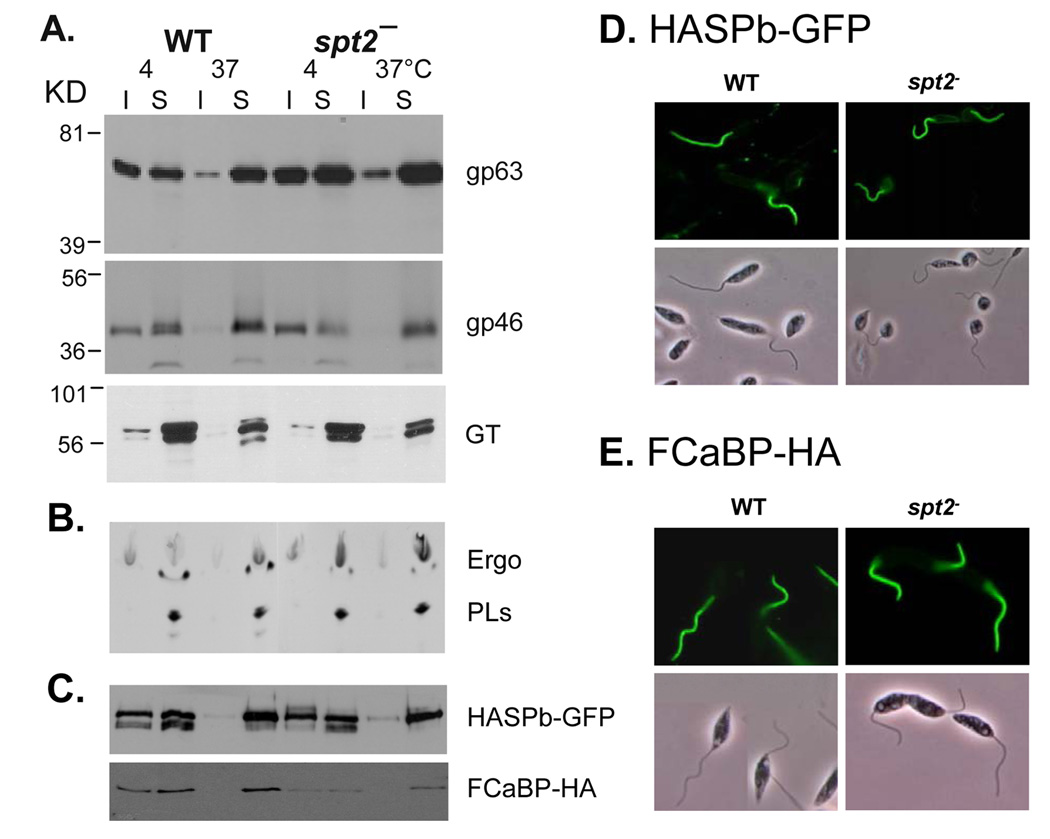
SL-null L. major generates normal DRM rafts. (A) TX100 insoluble (I) and soluble (S) fractions from log phase parasites (WT and spt2−) were analyzed by western-blot to assess the distribution of gp63, gp46, and glucose transporter (GT). (B) Log phase WT and spt2− parasites were labeled with [1,2]14C-acetate and extracted with 1% TX100 at 4 °C or 37 °C. Detergent insoluble materials were separated from soluble materials by centrifugation. Lipids from both insoluble and soluble fractions were extracted and analyzed by thin layer chromatography. Ergo: ergosterol; PLs: phospholipids. (C) WT and spt2− parasites with episomally expressed HASPB-GFP or FCaBP-HA were subjected to detergent extraction and western-blot analysis as described in (A), using anti-GFP and anti-HA antibodies. In addition, these parasites were subjected to fluorescence microscopy, as shown in D (GFP epifluorescence) and E (primary antibody: anti-HA mAb; secondary antibody: FITC-labeled goat-antimouse IgG). Leishmania culture, DRM isolation, and western-blot were performed as previously described [27]. To isolate metacyclics, spt2− mutants were grown to stationary phase in the presence of 500 µM of EtN as previously described [23].
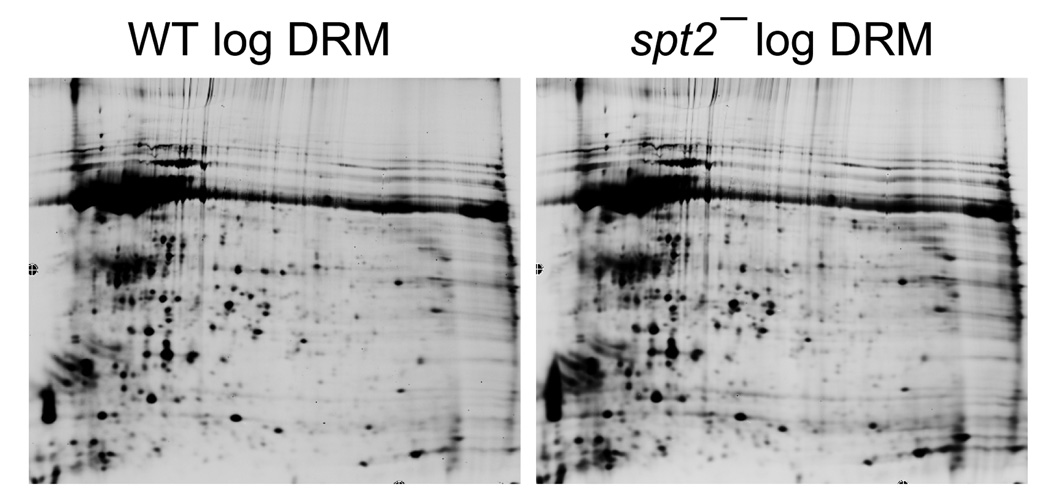
2D gel analysis of DRM fraction in L. major. DRM fractions were prepared from log phase WT and spt2− parasites as described [27] and subjected to 2D gel electrophoresis. Among all detectable spots, only ~1.2% showed greater than 2.5-fold variation between WT and spt2−.
Similar articles
-
Redirection of sphingolipid metabolism toward de novo synthesis of ethanolamine in Leishmania.EMBO J. 2007 Feb 21;26(4):1094-104. doi: 10.1038/sj.emboj.7601565. Epub 2007 Feb 8. EMBO J. 2007. PMID: 17290222 Free PMC article.
-
Sphingolipids in parasitic protozoa.Adv Exp Med Biol. 2010;688:238-48. doi: 10.1007/978-1-4419-6741-1_17. Adv Exp Med Biol. 2010. PMID: 20919659 Free PMC article. Review.
-
Complex Interplay between Sphingolipid and Sterol Metabolism Revealed by Perturbations to the Leishmania Metabolome Caused by Miltefosine.Antimicrob Agents Chemother. 2018 Apr 26;62(5):e02095-17. doi: 10.1128/AAC.02095-17. Print 2018 May. Antimicrob Agents Chemother. 2018. PMID: 29463533 Free PMC article.
-
Leishmania salvage and remodelling of host sphingolipids in amastigote survival and acidocalcisome biogenesis.Mol Microbiol. 2005 Mar;55(5):1566-78. doi: 10.1111/j.1365-2958.2005.04493.x. Mol Microbiol. 2005. PMID: 15720561 Free PMC article.
-
Phospholipids and Sphingolipids in Osteoarthritis.Biomolecules. 2025 Feb 8;15(2):250. doi: 10.3390/biom15020250. Biomolecules. 2025. PMID: 40001553 Free PMC article. Review.
Cited by
-
Phosphatidylcholine synthesis through cholinephosphate cytidylyltransferase is dispensable in Leishmania major.Sci Rep. 2019 May 20;9(1):7602. doi: 10.1038/s41598-019-44086-6. Sci Rep. 2019. PMID: 31110206 Free PMC article.
-
Expanded genome-wide comparisons give novel insights into population structure and genetic heterogeneity of Leishmania tropica complex.PLoS Negl Trop Dis. 2020 Sep 18;14(9):e0008684. doi: 10.1371/journal.pntd.0008684. eCollection 2020 Sep. PLoS Negl Trop Dis. 2020. PMID: 32946436 Free PMC article.
-
In vitro infectivity and differential gene expression of Leishmania infantum metacyclic promastigotes: negative selection with peanut agglutinin in culture versus isolation from the stomodeal valve of Phlebotomus perniciosus.BMC Genomics. 2016 May 20;17:375. doi: 10.1186/s12864-016-2672-8. BMC Genomics. 2016. PMID: 27206922 Free PMC article.
-
BluePort: a platform to study the eosinophilic response of mice to the bite of a vector of Leishmania parasites, Lutzomyia longipalpis sand flies.PLoS One. 2010 Oct 27;5(10):e13546. doi: 10.1371/journal.pone.0013546. PLoS One. 2010. PMID: 21048957 Free PMC article.
-
The Glycerol-3-Phosphate Acyltransferase TbGAT is Dispensable for Viability and the Synthesis of Glycerolipids in Trypanosoma brucei.J Eukaryot Microbiol. 2016 Sep;63(5):598-609. doi: 10.1111/jeu.12309. Epub 2016 Mar 8. J Eukaryot Microbiol. 2016. PMID: 26909872 Free PMC article.
References
-
- Majerus PW, et al. The metabolism of phosphoinositide-derived messenger molecules. Science. 1986;234:1519–1526. - PubMed
-
- Chakraborti S. Phospholipase A(2) isoforms: a perspective. Cell Signal. 2003;15:637–665. - PubMed
-
- Divecha N, Irvine RF. Phospholipid signaling. Cell. 1995;80:269–278. - PubMed
-
- Coppolino MG, et al. Inhibition of phosphatidylinositol-4-phosphate 5-kinase Ialpha impairs localized actin remodeling and suppresses phagocytosis. J Biol Chem. 2002;277:43849–43857. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
