

DNA supercoiling and its role in DNA decatenation and unknotting
- PMID: 20026582
- PMCID: PMC2853108
- DOI: 10.1093/nar/gkp1161
DNA supercoiling and its role in DNA decatenation and unknotting
Abstract
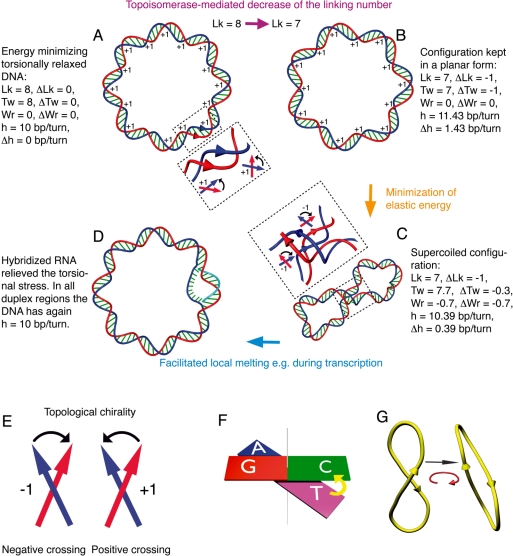
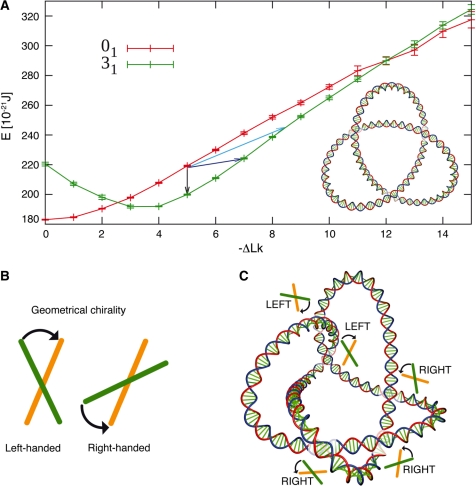
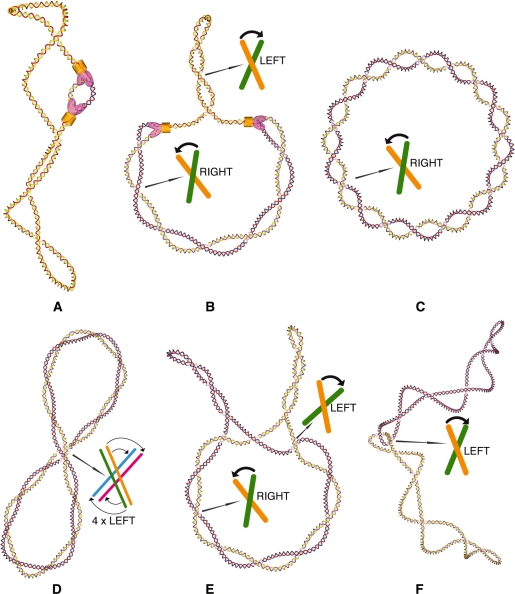
Chromosomal and plasmid DNA molecules in bacterial cells are maintained under torsional tension and are therefore supercoiled. With the exception of extreme thermophiles, supercoiling has a negative sign, which means that the torsional tension diminishes the DNA helicity and facilitates strand separation. In consequence, negative supercoiling aids such processes as DNA replication or transcription that require global- or local-strand separation. In extreme thermophiles, DNA is positively supercoiled which protects it from thermal denaturation. While the role of DNA supercoiling connected to the control of DNA stability, is thoroughly researched and subject of many reviews, a less known role of DNA supercoiling emerges and consists of aiding DNA topoisomerases in DNA decatenation and unknotting. Although DNA catenanes are natural intermediates in the process of DNA replication of circular DNA molecules, it is necessary that they become very efficiently decatenated, as otherwise the segregation of freshly replicated DNA molecules would be blocked. DNA knots arise as by-products of topoisomerase-mediated intramolecular passages that are needed to facilitate general DNA metabolism, including DNA replication, transcription or recombination. The formed knots are, however, very harmful for cells if not removed efficiently. Here, we overview the role of DNA supercoiling in DNA unknotting and decatenation.
Figures

Similar articles
-
Positive supercoiling of mitotic DNA drives decatenation by topoisomerase II in eukaryotes.Science. 2011 Mar 11;331(6022):1328-32. doi: 10.1126/science.1201538. Science. 2011. PMID: 21393545
-
Generation of supercoils in nicked and gapped DNA drives DNA unknotting and postreplicative decatenation.Nucleic Acids Res. 2015 Sep 3;43(15):7229-36. doi: 10.1093/nar/gkv683. Epub 2015 Jul 6. Nucleic Acids Res. 2015. PMID: 26150424 Free PMC article.
-
Topo IV is the topoisomerase that knots and unknots sister duplexes during DNA replication.Nucleic Acids Res. 2012 Apr;40(8):3563-73. doi: 10.1093/nar/gkr1237. Epub 2011 Dec 19. Nucleic Acids Res. 2012. PMID: 22187153 Free PMC article.
-
Closing the DNA replication cycle: from simple circular molecules to supercoiled and knotted DNA catenanes.Nucleic Acids Res. 2019 Aug 22;47(14):7182-7198. doi: 10.1093/nar/gkz586. Nucleic Acids Res. 2019. PMID: 31276584 Free PMC article. Review.
-
[DNA supercoiling and topoisomerases in Escherichia coli].Rev Latinoam Microbiol. 1995 Jul-Sep;37(3):291-304. Rev Latinoam Microbiol. 1995. PMID: 8850348 Review. Spanish.
Cited by
-
Interplay between type 1A topoisomerases and gyrase in chromosome segregation in Escherichia coli.J Bacteriol. 2013 Apr;195(8):1758-68. doi: 10.1128/JB.02001-12. Epub 2013 Feb 8. J Bacteriol. 2013. PMID: 23396913 Free PMC article.
-
Knot Factories with Helical Geometry Enhance Knotting and Induce Handedness to Knots.Polymers (Basel). 2022 Oct 7;14(19):4201. doi: 10.3390/polym14194201. Polymers (Basel). 2022. PMID: 36236148 Free PMC article.
-
Mutagenesis of Intrinsically Disordered Domain Impacts Topoisomerase IIα Catalytic Activity.Int J Mol Sci. 2025 Apr 11;26(8):3604. doi: 10.3390/ijms26083604. Int J Mol Sci. 2025. PMID: 40332112 Free PMC article.
-
Direct Evidence for the Formation of Precatenanes during DNA Replication.J Biol Chem. 2015 May 29;290(22):13725-35. doi: 10.1074/jbc.M115.642272. Epub 2015 Mar 31. J Biol Chem. 2015. PMID: 25829493 Free PMC article.
-
The polypyrimidine/polypurine motif in the mouse mu opioid receptor gene promoter is a supercoiling-regulatory element.Gene. 2011 Nov 1;487(1):52-61. doi: 10.1016/j.gene.2011.07.026. Epub 2011 Jul 31. Gene. 2011. PMID: 21839154 Free PMC article.
References
-
- Cloutier TE, Widom J. Spontaneous sharp bending of double-stranded DNA. Mol. Cell. 2004;14:355–362. - PubMed
-
- Vologodskii AV, Levene SD, Klenin KV, Frank-Kamenetskii M, Cozzarelli NR. Conformational and thermodynamic properties of supercoiled DNA. J. Mol. Biol. 1992;227:1224–1243. - PubMed
-
- Strick TR, Allemand JF, Bensimon D, Bensimon A, Croquette V. The elasticity of a single supercoiled DNA molecule. Science. 1996;271:1835–1837. - PubMed
