

Dynamic changes in the subcellular distribution of Gpd1p in response to cell stress
- PMID: 20026609
- PMCID: PMC2825468
- DOI: 10.1074/jbc.M109.058552
Dynamic changes in the subcellular distribution of Gpd1p in response to cell stress
Abstract
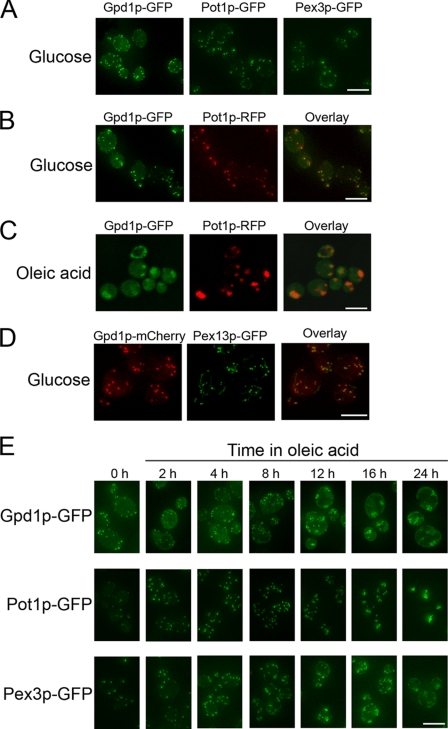
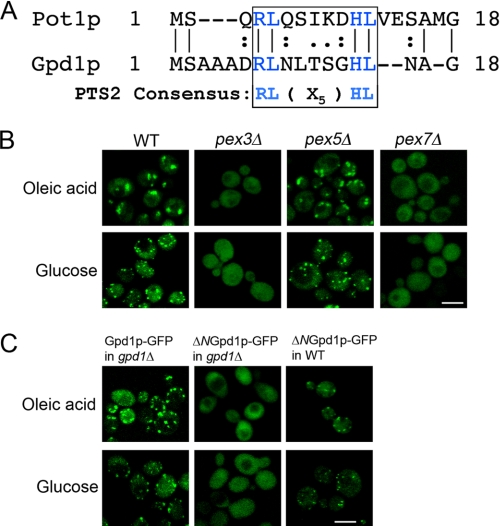
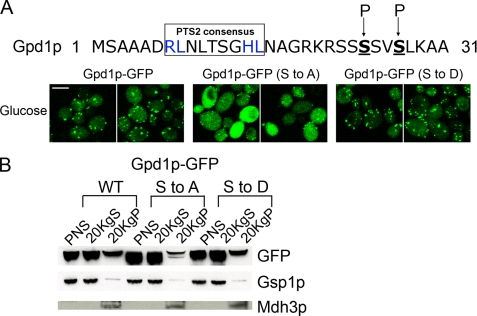
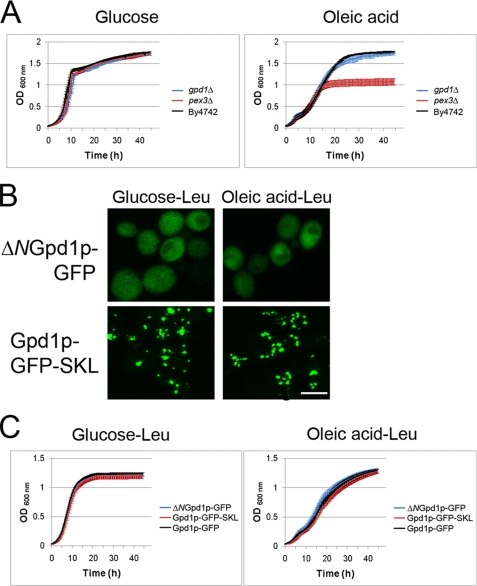
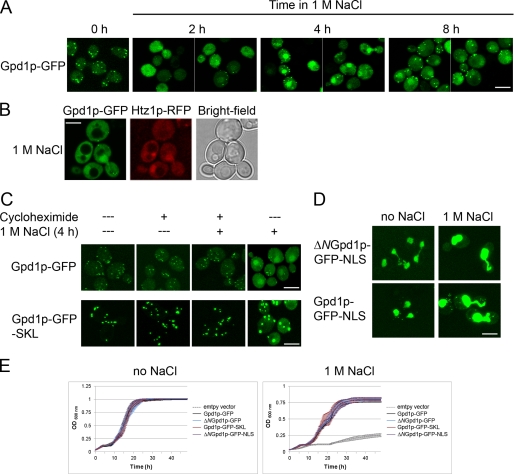
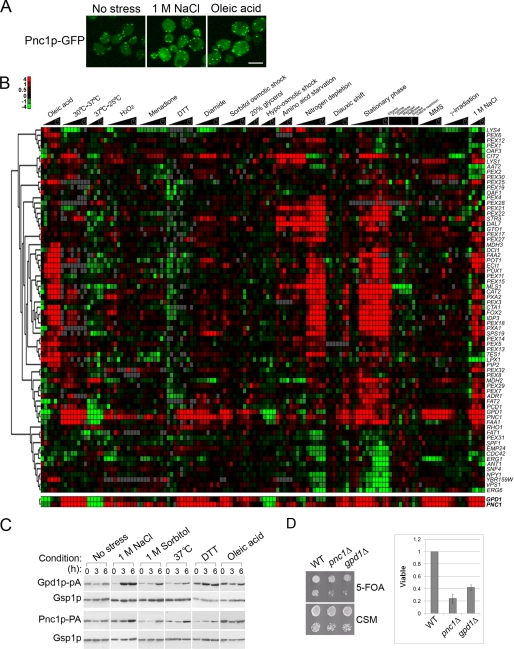
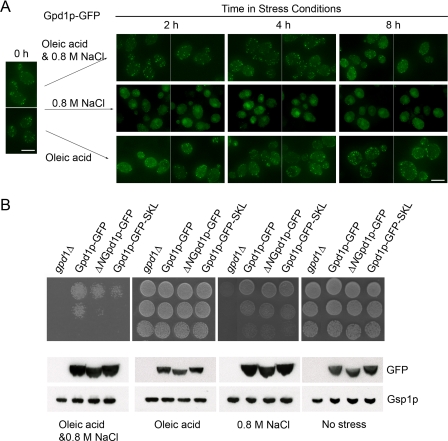
Gpd1p is a cytosolic NAD(+)-dependent glycerol 3-phosphate dehydrogenase that also localizes to peroxisomes and plays an essential role in the cellular response to osmotic stress and a role in redox balance. Here, we show that Gpd1p is directed to peroxisomes by virtue of an N-terminal type 2 peroxisomal targeting signal (PTS2) in a Pex7p-dependent manner. Significantly, localization of Gpd1p to peroxisomes is dependent on the metabolic status of cells and the phosphorylation of aminoacyl residues adjacent to the targeting signal. Exposure of cells to osmotic stress induces changes in the subcellular distribution of Gpd1p to the cytosol and nucleus. This behavior is similar to Pnc1p, which is coordinately expressed with Gpd1p, and under conditions of cell stress changes its subcellular distribution from peroxisomes to the nucleus where it mediates chromatin silencing. Although peroxisomes are necessary for the beta-oxidation of fatty acids in yeast, the localization of Gpd1p to peroxisomes is not. Rather, shifts in the distribution of Gpd1p to different cellular compartments in response to changing cellular status suggests a role for Gpd1p in the spatial regulation of redox potential, a process critical to cell survival, especially under the complex stress conditions expected to occur in the wild.
Figures
Similar articles
-
Role of Pex21p for Piggyback Import of Gpd1p and Pnc1p into Peroxisomes of Saccharomyces cerevisiae.J Biol Chem. 2015 Oct 16;290(42):25333-42. doi: 10.1074/jbc.M115.653451. Epub 2015 Aug 15. J Biol Chem. 2015. PMID: 26276932 Free PMC article.
-
Distinct intracellular localization of Gpd1p and Gpd2p, the two yeast isoforms of NAD+-dependent glycerol-3-phosphate dehydrogenase, explains their different contributions to redox-driven glycerol production.J Biol Chem. 2004 Sep 17;279(38):39677-85. doi: 10.1074/jbc.M403310200. Epub 2004 Jun 21. J Biol Chem. 2004. PMID: 15210723
-
The cytosolic peroxisome-targeting signal (PTS)-receptors, Pex7p and Pex5pL, are sufficient to transport PTS2 proteins to peroxisomes.Biochim Biophys Acta Mol Cell Res. 2019 Mar;1866(3):441-449. doi: 10.1016/j.bbamcr.2018.10.006. Epub 2018 Oct 5. Biochim Biophys Acta Mol Cell Res. 2019. PMID: 30296498
-
Fatty Acid Oxidation in Peroxisomes: Enzymology, Metabolic Crosstalk with Other Organelles and Peroxisomal Disorders.Adv Exp Med Biol. 2020;1299:55-70. doi: 10.1007/978-3-030-60204-8_5. Adv Exp Med Biol. 2020. PMID: 33417207 Review.
-
The type-2 peroxisomal targeting signal.Biochim Biophys Acta Mol Cell Res. 2020 Feb;1867(2):118609. doi: 10.1016/j.bbamcr.2019.118609. Epub 2019 Nov 18. Biochim Biophys Acta Mol Cell Res. 2020. PMID: 31751594 Review.
Cited by
-
ESCRT-III is required for scissioning new peroxisomes from the endoplasmic reticulum.J Cell Biol. 2018 Jun 4;217(6):2087-2102. doi: 10.1083/jcb.201706044. Epub 2018 Mar 27. J Cell Biol. 2018. PMID: 29588378 Free PMC article.
-
Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat--store 'em up or burn 'em down.Genetics. 2013 Jan;193(1):1-50. doi: 10.1534/genetics.112.143362. Genetics. 2013. PMID: 23275493 Free PMC article. Review.
-
Identification of Arabidopsis Protein Kinases That Harbor Functional Type 1 Peroxisomal Targeting Signals.Front Cell Dev Biol. 2022 Feb 15;10:745883. doi: 10.3389/fcell.2022.745883. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35242755 Free PMC article.
-
Peroxisomal Import Reduces the Proapoptotic Activity of Deubiquitinating Enzyme USP2.PLoS One. 2015 Oct 20;10(10):e0140685. doi: 10.1371/journal.pone.0140685. eCollection 2015. PLoS One. 2015. PMID: 26484888 Free PMC article.
-
TOR and heat shock response pathways regulate peroxisome biogenesis during proteotoxic stress.bioRxiv [Preprint]. 2025 Jan 2:2024.12.31.630809. doi: 10.1101/2024.12.31.630809. bioRxiv. 2025. PMID: 40093121 Free PMC article. Preprint.
References
-
- Larsson K., Ansell R., Eriksson P., Adler L. (1993) Mol. Microbiol. 10, 1101–1111 - PubMed
-
- Larsson C., Påhlman I. L., Ansell R., Rigoulet M., Adler L., Gustafsson L. (1998) Yeast 14, 347–357 - PubMed
-
- Aguilera J., Rodríguez-Vargas S., Prieto J. A. (2005) Mol. Microbiol. 56, 228–239 - PubMed
-
- Phillips S. A., Thornalley P. J. (1993) Eur. J. Biochem. 212, 101–105 - PubMed
-
- Vander Jagt D. L., Robinson B., Taylor K. K., Hunsaker L. A. (1992) J. Biol. Chem. 267, 4364–4369 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
