

A neo-sex chromosome that drives postzygotic sex determination in the hessian fly (Mayetiola destructor)
- PMID: 20026681
- PMCID: PMC2845344
- DOI: 10.1534/genetics.109.108589
A neo-sex chromosome that drives postzygotic sex determination in the hessian fly (Mayetiola destructor)
Abstract
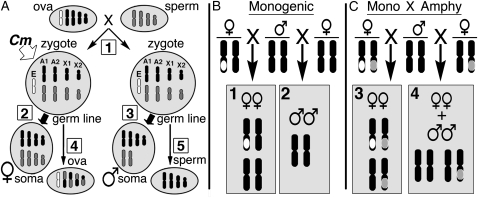
Two nonoverlapping autosomal inversions defined unusual neo-sex chromosomes in the Hessian fly (Mayetiola destructor). Like other neo-sex chromosomes, these were normally heterozygous, present only in one sex, and suppressed recombination around a sex-determining master switch. Their unusual properties originated from the anomalous Hessian fly sex determination system in which postzygotic chromosome elimination is used to establish the sex-determining karyotypes. This system permitted the evolution of a master switch (Chromosome maintenance, Cm) that acts maternally. All of the offspring of females that carry Cm-associated neo-sex chromosomes attain a female-determining somatic karyotype and develop as females. Thus, the chromosomes act as maternal effect neo-W's, or W-prime (W') chromosomes, where ZW' females mate with ZZ males to engender female-producing (ZW') and male-producing (ZZ) females in equal numbers. Genetic mapping and physical mapping identified the inversions. Their distribution was determined in nine populations. Experimental matings established the association of the inversions with Cm and measured their recombination suppression. The inversions are the functional equivalent of the sciarid X-prime chromosomes. We speculate that W' chromosomes exist in a variety of species that produce unisexual broods.
Figures




Similar articles
-
Evidence for an unusual ZW/ZW'/ZZ sex-chromosome system in Scardinius erythrophthalmus (Pisces, Cyprinidae), as detected by cytogenetic and H-Y antigen analyses.Cytogenet Cell Genet. 1995;71(4):356-62. doi: 10.1159/000134139. Cytogenet Cell Genet. 1995. PMID: 8521724
-
Recent Evolution of a Maternally Acting Sex-Determining Supergene in a Fly with Single-Sex Broods.Mol Biol Evol. 2023 Jul 5;40(7):msad148. doi: 10.1093/molbev/msad148. Mol Biol Evol. 2023. PMID: 37352554 Free PMC article.
-
Reconstruction of female heterogamety from admixture of XX-XY and ZZ-ZW sex-chromosome systems within a frog species.Mol Ecol. 2018 Oct;27(20):4078-4089. doi: 10.1111/mec.14831. Epub 2018 Sep 4. Mol Ecol. 2018. PMID: 30086193
-
The Cause-Effect Model of Master Sex Determination Gene Acquisition and the Evolution of Sex Chromosomes.Int J Mol Sci. 2025 Apr 1;26(7):3282. doi: 10.3390/ijms26073282. Int J Mol Sci. 2025. PMID: 40244140 Free PMC article. Review.
-
Avian sex, sex chromosomes, and dosage compensation in the age of genomics.Chromosome Res. 2014 Apr;22(1):45-57. doi: 10.1007/s10577-014-9409-9. Chromosome Res. 2014. PMID: 24599719 Review.
Cited by
-
Male death resulting from hybridization between subspecies of the gypsy moth, Lymantria dispar.Heredity (Edinb). 2011 Apr;106(4):603-13. doi: 10.1038/hdy.2010.92. Epub 2010 Jul 14. Heredity (Edinb). 2011. PMID: 20628417 Free PMC article.
-
Deep sequencing and genome-wide analysis reveals the expansion of MicroRNA genes in the gall midge Mayetiola destructor.BMC Genomics. 2013 Mar 18;14:187. doi: 10.1186/1471-2164-14-187. BMC Genomics. 2013. PMID: 23496979 Free PMC article.
-
Use of microsatellite and SNP markers for biotype characterization in Hessian fly.J Insect Sci. 2015 Nov 5;15(1):158. doi: 10.1093/jisesa/iev138. Print 2015. J Insect Sci. 2015. PMID: 26543089 Free PMC article.
-
Non-Mendelian transmission of X chromosomes: mechanisms and impact on sex ratios and population dynamics in different breeding systems.Biochem Soc Trans. 2024 Aug 28;52(4):1777-1784. doi: 10.1042/BST20231411. Biochem Soc Trans. 2024. PMID: 39149984 Free PMC article. Review.
-
An equal sex ratio followed by differential sex mortality causes overestimation of females in gall midges: no evidence for sex ratio regulation.Naturwissenschaften. 2012 Jun;99(6):493-9. doi: 10.1007/s00114-012-0925-1. Epub 2012 May 29. Naturwissenschaften. 2012. PMID: 22643882
References
-
- Albertson, D. G., O. C. Nwaorgu and J. E. Sulston, 1979. Chromatin diminution and a chromosomal mechanism of sexual differentiation in Strongyloides papillosus. Chromosoma 75 75–87. - PubMed
-
- Bachtrog, D., 2005. Adaptation shapes patterns of genome evolution on sexual and asexual chromosomes in Drosophila. Nat. Genet. 34 215–219. - PubMed
-
- Bell, G., 1982. The Masterpiece of Nature: The Evolution and Genetics of Sexuality. University of California Press, Berkeley, CA/Los Angeles.
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
