

Human host factors required for influenza virus replication
- PMID: 20027183
- PMCID: PMC2862546
- DOI: 10.1038/nature08699
Human host factors required for influenza virus replication
Abstract
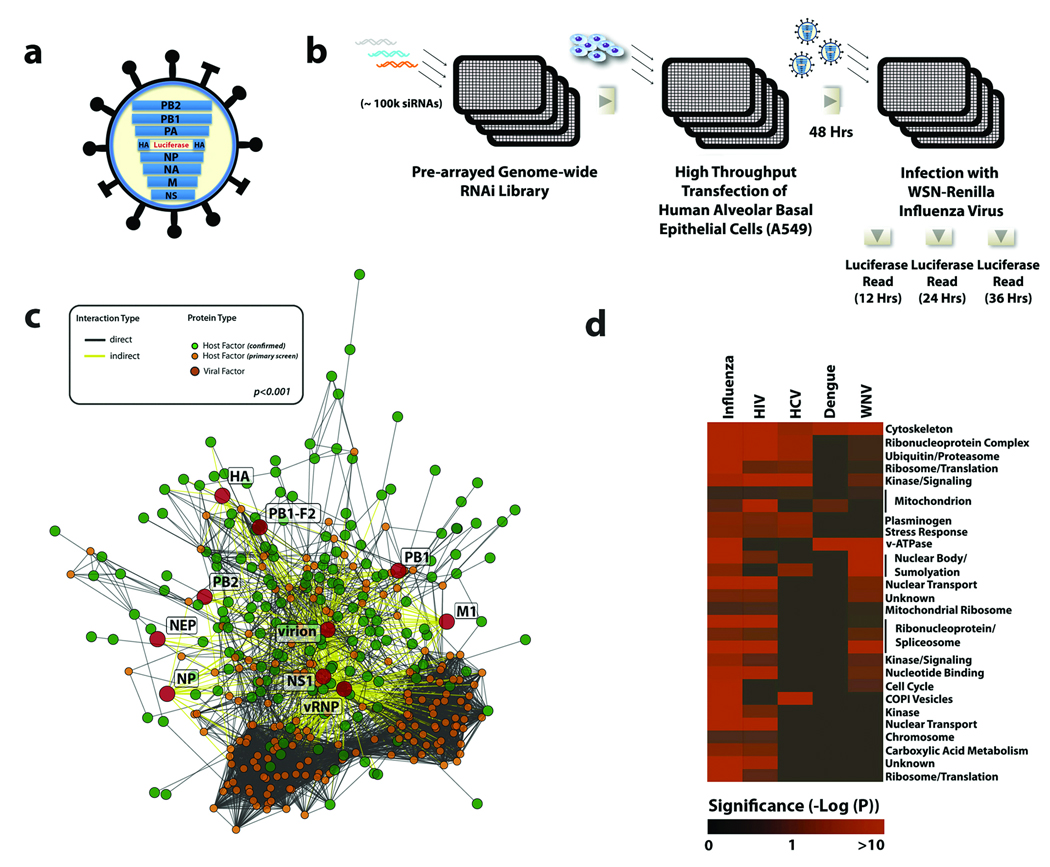
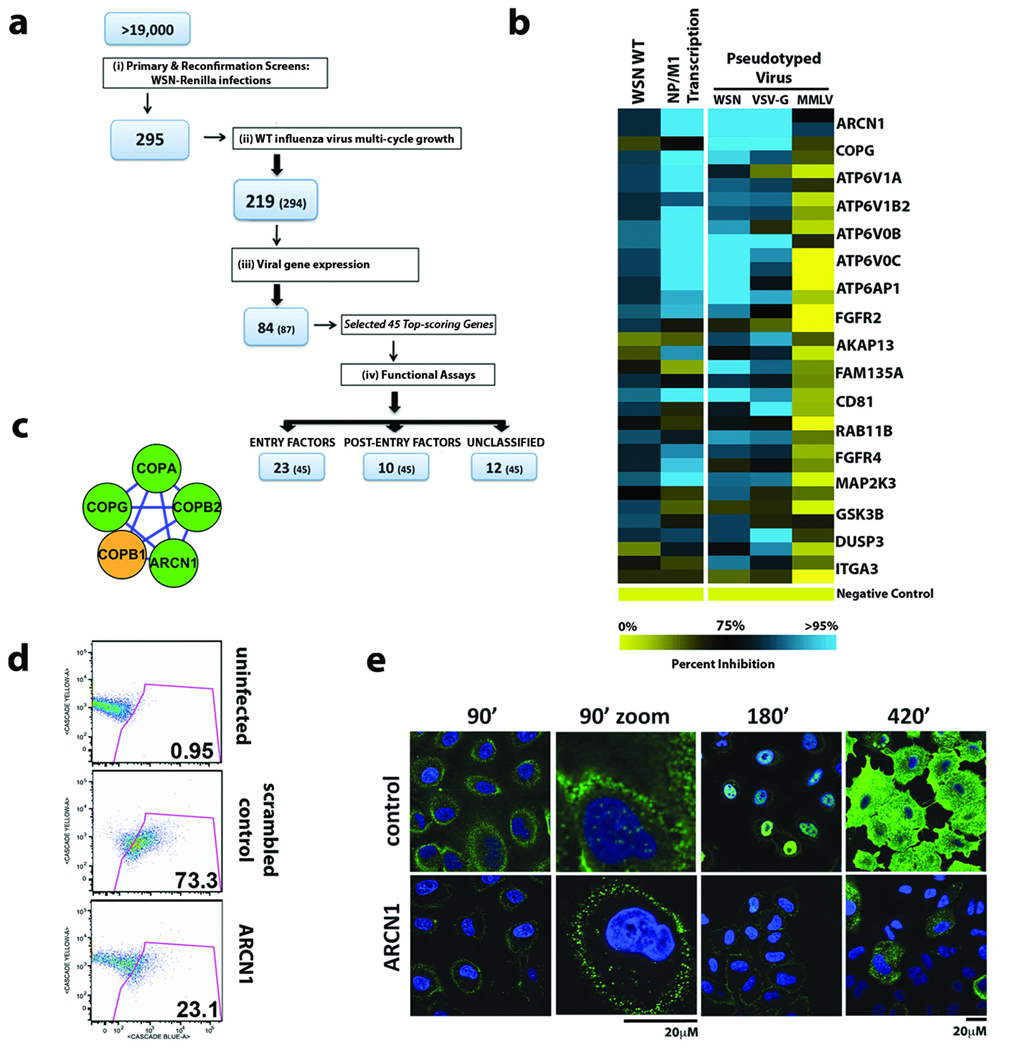
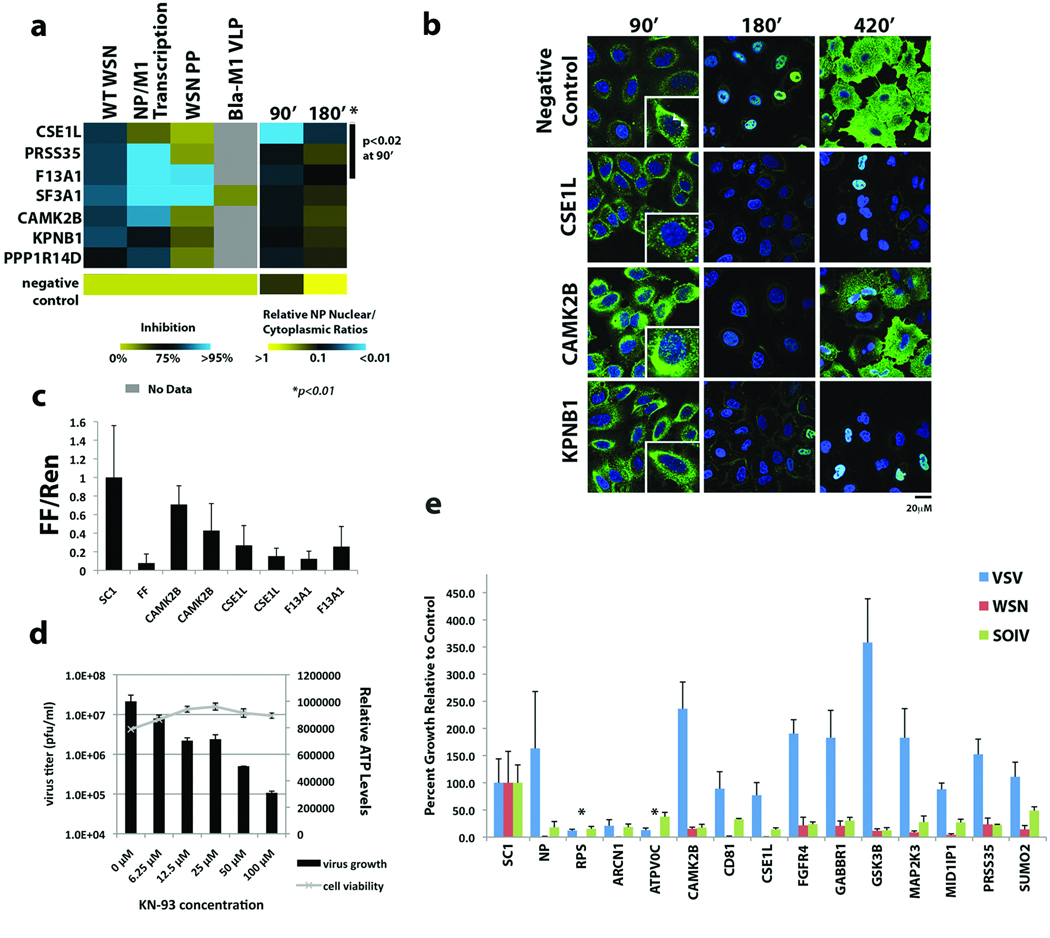
Influenza A virus is an RNA virus that encodes up to 11 proteins and this small coding capacity demands that the virus use the host cellular machinery for many aspects of its life cycle. Knowledge of these host cell requirements not only informs us of the molecular pathways exploited by the virus but also provides further targets that could be pursued for antiviral drug development. Here we use an integrative systems approach, based on genome-wide RNA interference screening, to identify 295 cellular cofactors required for early-stage influenza virus replication. Within this group, those involved in kinase-regulated signalling, ubiquitination and phosphatase activity are the most highly enriched, and 181 factors assemble into a highly significant host-pathogen interaction network. Moreover, 219 of the 295 factors were confirmed to be required for efficient wild-type influenza virus growth, and further analysis of a subset of genes showed 23 factors necessary for viral entry, including members of the vacuolar ATPase (vATPase) and COPI-protein families, fibroblast growth factor receptor (FGFR) proteins, and glycogen synthase kinase 3 (GSK3)-beta. Furthermore, 10 proteins were confirmed to be involved in post-entry steps of influenza virus replication. These include nuclear import components, proteases, and the calcium/calmodulin-dependent protein kinase (CaM kinase) IIbeta (CAMK2B). Notably, growth of swine-origin H1N1 influenza virus is also dependent on the identified host factors, and we show that small molecule inhibitors of several factors, including vATPase and CAMK2B, antagonize influenza virus replication.
Figures
References
-
- Palese P, Shaw ML. Orthomyxoviridae: The Viruses and Their Replication. In: Knipe DM, Howley PM, editors. Fields Virology. 5th Edition. Vol. 2. Philadelphia: Lippincott Williams & Wilkins; 2007. pp. 1647–1689.
-
- Wang TT, Palese P. Unraveling the mystery of swine influenza virus. Cell. 2009;137:983–985. - PubMed
-
- Brass AL, et al. Identification of host proteins required for HIV infection through a functional genomic screen. Science. 2008;319:921–926. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- HHSN272200900032C/AI/NIAID NIH HHS/United States
- U54 AI057159/AI/NIAID NIH HHS/United States
- F32 AI081428/AI/NIAID NIH HHS/United States
- R21 AI083673/AI/NIAID NIH HHS/United States
- 1 P01 AI058113/AI/NIAID NIH HHS/United States
- U54 AI057158/AI/NIAID NIH HHS/United States
- 1 T32 AI07647/AI/NIAID NIH HHS/United States
- T32 AI007647/AI/NIAID NIH HHS/United States
- 1F32AI081428/AI/NIAID NIH HHS/United States
- U01 AI1074539/AI/NIAID NIH HHS/United States
- 1R21AI083673/AI/NIAID NIH HHS/United States
- 5R24 CA095823-04/CA/NCI NIH HHS/United States
- 1 S10 RR0 9145-01/RR/NCRR NIH HHS/United States
- U01 AI074539/AI/NIAID NIH HHS/United States
- R24 CA095823/CA/NCI NIH HHS/United States
- T32 GM007280/GM/NIGMS NIH HHS/United States
- HHSN266200700010C/AI/NIAID NIH HHS/United States
- P01 AI058113/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
