

Opposite effects of low and high doses of Abeta42 on electrical network and neuronal excitability in the rat prefrontal cortex
- PMID: 20027222
- PMCID: PMC2791225
- DOI: 10.1371/journal.pone.0008366
Opposite effects of low and high doses of Abeta42 on electrical network and neuronal excitability in the rat prefrontal cortex
Abstract
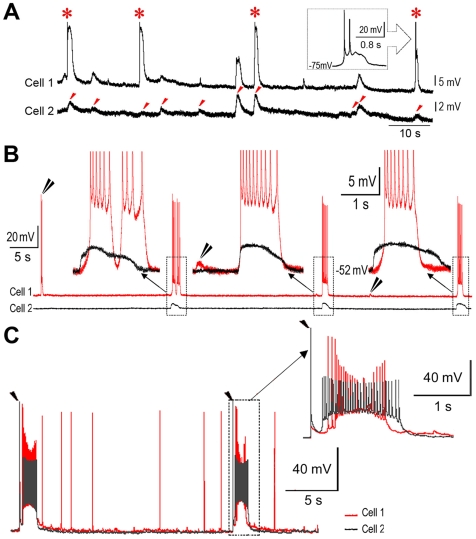
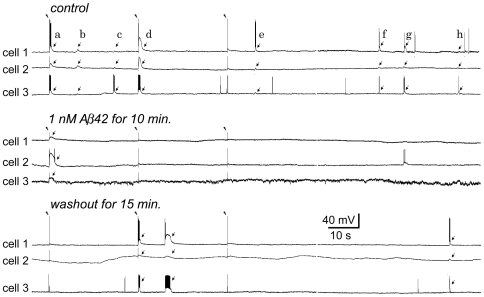
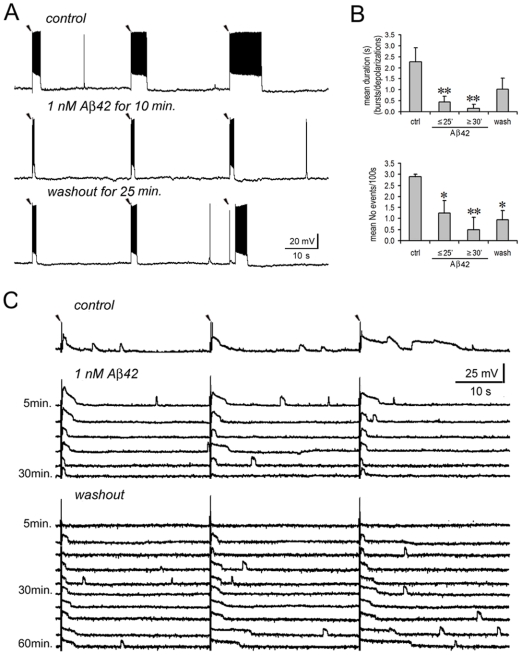
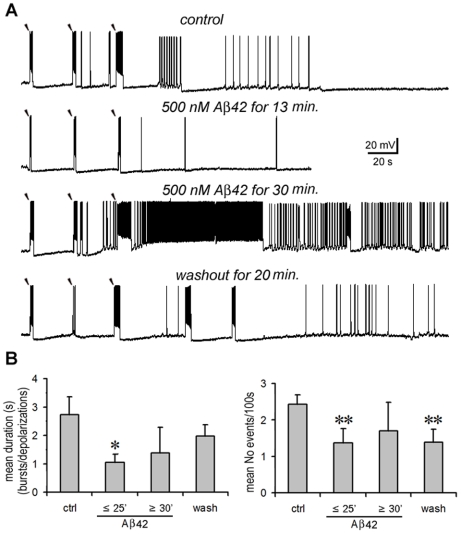
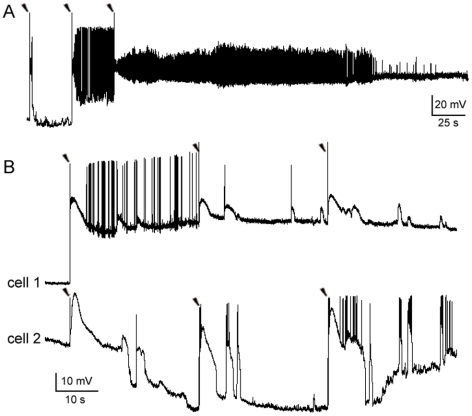
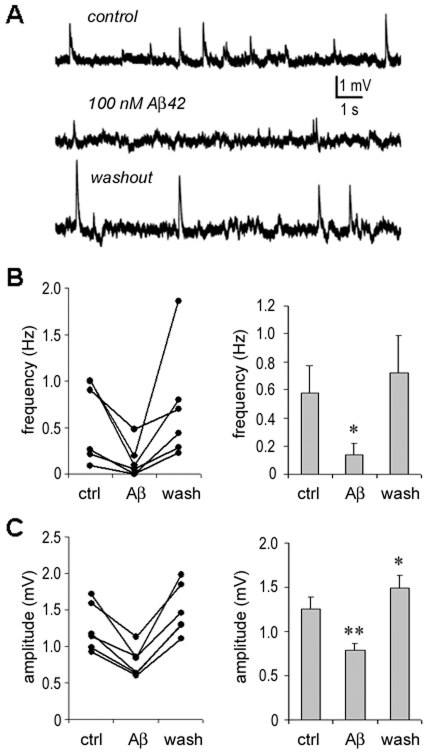
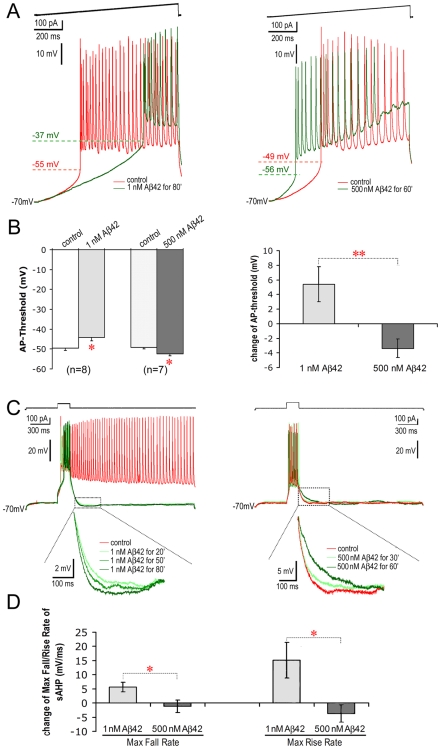
Changes in neuronal synchronization have been found in patients and animal models of Alzheimer's disease (AD). Synchronized behaviors within neuronal networks are important to such complex cognitive processes as working memory. The mechanisms behind these changes are not understood but may involve the action of soluble beta-amyloid (Abeta) on electrical networks. In order to determine if Abeta can induce changes in neuronal synchronization, the activities of pyramidal neurons were recorded in rat prefrontal cortical (PFC) slices under calcium-free conditions using multi-neuron patch clamp technique. Electrical network activities and synchronization among neurons were significantly inhibited by low dose Abeta42 (1 nM) and initially by high dose Abeta42 (500 nM). However, prolonged application of high dose Abeta42 resulted in network activation and tonic firing. Underlying these observations, we discovered that prolonged application of low and high doses of Abeta42 induced opposite changes in action potential (AP)-threshold and after-hyperpolarization (AHP) of neurons. Accordingly, low dose Abeta42 significantly increased the AP-threshold and deepened the AHP, making neurons less excitable. In contrast, high dose Abeta42 significantly reduced the AP-threshold and shallowed the AHP, making neurons more excitable. These results support a model that low dose Abeta42 released into the interstitium has a physiologic feedback role to dampen electrical network activity by reducing neuronal excitability. Higher concentrations of Abeta42 over time promote supra-synchronization between individual neurons by increasing their excitability. The latter may disrupt frontal-based cognitive processing and in some cases lead to epileptiform discharges.
Conflict of interest statement
Figures
Similar articles
-
Intrinsic excitability changes induced by acute treatment of hippocampal CA1 pyramidal neurons with exogenous amyloid β peptide.Hippocampus. 2015 Jul;25(7):786-97. doi: 10.1002/hipo.22403. Epub 2015 Mar 25. Hippocampus. 2015. PMID: 25515596 Free PMC article.
-
Alterations in CA1 pyramidal neuronal intrinsic excitability mediated by Ih channel currents in a rat model of amyloid beta pathology.Neuroscience. 2015 Oct 1;305:279-92. doi: 10.1016/j.neuroscience.2015.07.087. Epub 2015 Aug 5. Neuroscience. 2015. PMID: 26254243
-
Dopamine increases excitability of pyramidal neurons in primate prefrontal cortex.J Neurophysiol. 2000 Dec;84(6):2799-809. doi: 10.1152/jn.2000.84.6.2799. J Neurophysiol. 2000. PMID: 11110810
-
Amyloid-β-induced action potential desynchronization and degradation of hippocampal gamma oscillations is prevented by interference with peptide conformation change and aggregation.J Neurosci. 2014 Aug 20;34(34):11416-25. doi: 10.1523/JNEUROSCI.1195-14.2014. J Neurosci. 2014. PMID: 25143621 Free PMC article.
-
Targeting the alpha 7 nicotinic acetylcholine receptor to reduce amyloid accumulation in Alzheimer's disease pyramidal neurons.Curr Pharm Des. 2006;12(6):677-84. doi: 10.2174/138161206775474224. Curr Pharm Des. 2006. PMID: 16472157 Review.
Cited by
-
Amyloid β Peptide-Induced Changes in Prefrontal Cortex Activity and Its Response to Hippocampal Input.Int J Pept. 2017;2017:7386809. doi: 10.1155/2017/7386809. Epub 2017 Jan 3. Int J Pept. 2017. PMID: 28127312 Free PMC article.
-
The dysregulation of intracellular calcium in Alzheimer disease.Cell Calcium. 2010 Feb;47(2):183-9. doi: 10.1016/j.ceca.2009.12.014. Epub 2010 Jan 18. Cell Calcium. 2010. PMID: 20080301 Free PMC article. Review.
-
Amyloid burden is associated with self-reported sleep in nondemented late middle-aged adults.Neurobiol Aging. 2015 Sep;36(9):2568-76. doi: 10.1016/j.neurobiolaging.2015.05.004. Epub 2015 May 14. Neurobiol Aging. 2015. PMID: 26059712 Free PMC article.
-
Physiological Aβ Concentrations Produce a More Biomimetic Representation of the Alzheimer's Disease Phenotype in iPSC Derived Human Neurons.ACS Chem Neurosci. 2018 Jul 18;9(7):1693-1701. doi: 10.1021/acschemneuro.8b00067. Epub 2018 May 22. ACS Chem Neurosci. 2018. PMID: 29746089 Free PMC article.
-
K(Ca)2 and k(ca)3 channels in learning and memory processes, and neurodegeneration.Front Pharmacol. 2012 Jun 11;3:107. doi: 10.3389/fphar.2012.00107. eCollection 2012. Front Pharmacol. 2012. PMID: 22701424 Free PMC article.
References
-
- Hormuzdi SG, Filippov MA, Mitropoulou G, Monyer H, Bruzzone R. Electrical synapses: a dynamic signaling system that shapes the activity of neuronal networks. Biochim Biophys Acta. 2004;1662:113–137. - PubMed
-
- Bennett MV, Zukin RS. Electrical coupling and neuronal synchronization in the Mammalian brain. Neuron. 2004;41:495–511. - PubMed
-
- Uhlhaas PJ, Singer W. Neural synchrony in brain disorders: relevance for cognitive dysfunctions and pathophysiology. Neuron. 2006;52:155–168. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
