

Representation sharpening can explain perceptual priming
- PMID: 20028230
- PMCID: PMC2862237
- DOI: 10.1162/neco.2009.04-09-999
Representation sharpening can explain perceptual priming
Abstract
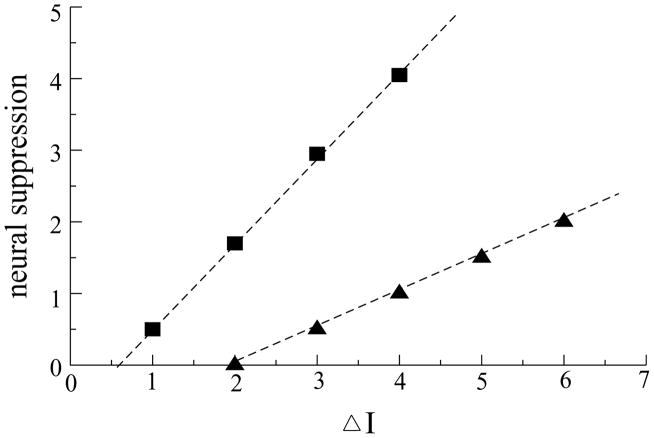
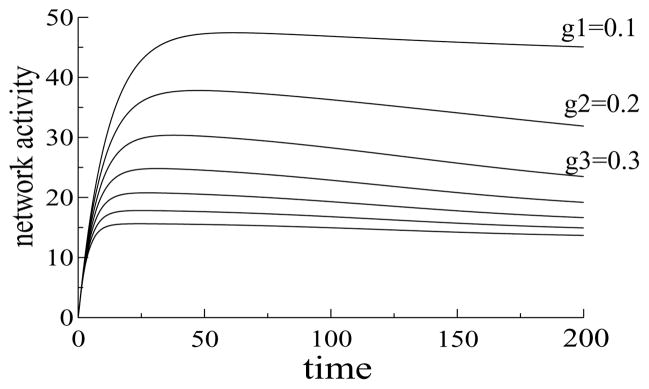
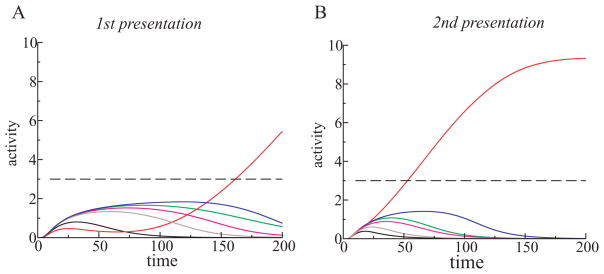
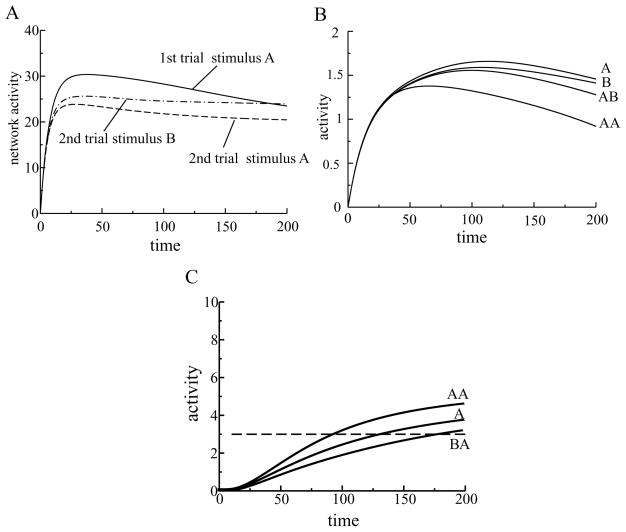
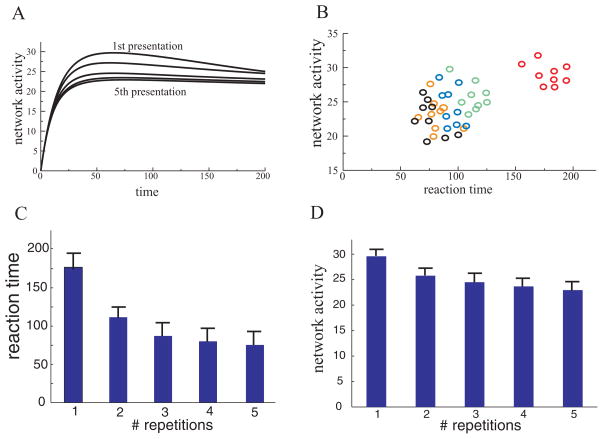
Perceiving and identifying an object is improved by prior exposure to the object. This perceptual priming phenomenon is accompanied by reduced neural activity. But whether suppression of neuronal activity with priming is responsible for the improvement in perception is unclear. To address this problem, we developed a rate-based network model of visual processing. In the model, decreased neural activity following priming was due to stimulus-specific sharpening of representations taking place in the early visual areas. Representation sharpening led to decreased interference of representations in higher visual areas that facilitated selection of one of the competing representations, thereby improving recognition. The model explained a wide range of psychophysical and physiological data observed in priming experiments, including antipriming phenomena, and predicted two functionally distinct stages of visual processing.
Figures
Similar articles
-
Perceptual priming leads to reduction of gamma frequency oscillations.Proc Natl Acad Sci U S A. 2010 Mar 23;107(12):5640-5. doi: 10.1073/pnas.0907525107. Epub 2010 Mar 8. Proc Natl Acad Sci U S A. 2010. PMID: 20212165 Free PMC article.
-
A Gradient of Sharpening Effects by Perceptual Prior across the Human Cortical Hierarchy.J Neurosci. 2021 Jan 6;41(1):167-178. doi: 10.1523/JNEUROSCI.2023-20.2020. Epub 2020 Nov 18. J Neurosci. 2021. PMID: 33208472 Free PMC article.
-
Unsupervised changes in core object recognition behavior are predicted by neural plasticity in inferior temporal cortex.Elife. 2021 Jun 11;10:e60830. doi: 10.7554/eLife.60830. Elife. 2021. PMID: 34114566 Free PMC article.
-
Single units and conscious vision.Philos Trans R Soc Lond B Biol Sci. 1998 Nov 29;353(1377):1801-18. doi: 10.1098/rstb.1998.0333. Philos Trans R Soc Lond B Biol Sci. 1998. PMID: 9854253 Free PMC article. Review.
-
Advances in visual perceptual learning and plasticity.Nat Rev Neurosci. 2010 Jan;11(1):53-60. doi: 10.1038/nrn2737. Epub 2009 Dec 2. Nat Rev Neurosci. 2010. PMID: 19953104 Free PMC article. Review.
Cited by
-
On the relationship between persistent delay activity, repetition enhancement and priming.Front Psychol. 2015 Jan 22;5:1590. doi: 10.3389/fpsyg.2014.01590. eCollection 2014. Front Psychol. 2015. PMID: 25657630 Free PMC article. Review.
-
Perceptual priming leads to reduction of gamma frequency oscillations.Proc Natl Acad Sci U S A. 2010 Mar 23;107(12):5640-5. doi: 10.1073/pnas.0907525107. Epub 2010 Mar 8. Proc Natl Acad Sci U S A. 2010. PMID: 20212165 Free PMC article.
-
Modulation of cross-frequency coupling by novel and repeated stimuli in the primate ventrolateral prefrontal cortex.Front Psychol. 2011 Sep 7;2:217. doi: 10.3389/fpsyg.2011.00217. eCollection 2011. Front Psychol. 2011. PMID: 21941517 Free PMC article.
-
Learning optimal strategies in complex environments.Proc Natl Acad Sci U S A. 2010 Nov 23;107(47):20151-2. doi: 10.1073/pnas.1014954107. Epub 2010 Nov 15. Proc Natl Acad Sci U S A. 2010. PMID: 21078996 Free PMC article. No abstract available.
References
-
- Blake R, Logothetis NK. Visual competition. Nat Rev Neurosci. 2001;3:1–11. - PubMed
-
- Bogacz R, Brown MW. Comparison of computational models of familiarity discrimination in the perirhinal cortex. Hippocampus. 2003;13:494–524. - PubMed
-
- Foeller E, Feldman DE. Synaptic basis for developmental plasticity in somatosensory cortex. Curr Opin Neurobiol. 2004;14:89–95. - PubMed
-
- Freedman DJ, Riesenhuber M, Poggio T, Miller EK. Experience-dependent sharpening of visual shape selectivity in inferior temporal cortex. Cereb Cortex. 2006;16:1631–1644. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources