

Splicing factor Spf30 assists exosome-mediated gene silencing in fission yeast
- PMID: 20028739
- PMCID: PMC2820897
- DOI: 10.1128/MCB.01317-09
Splicing factor Spf30 assists exosome-mediated gene silencing in fission yeast
Erratum in
-
Correction for Bernard et al., "Splicing Factor Spf30 Assists Exosome-Mediated Gene Silencing in Fission Yeast".Mol Cell Biol. 2017 May 16;37(11):e00115-17. doi: 10.1128/MCB.00115-17. Print 2017 Jun 1. Mol Cell Biol. 2017. PMID: 28512211 Free PMC article. No abstract available.
Abstract
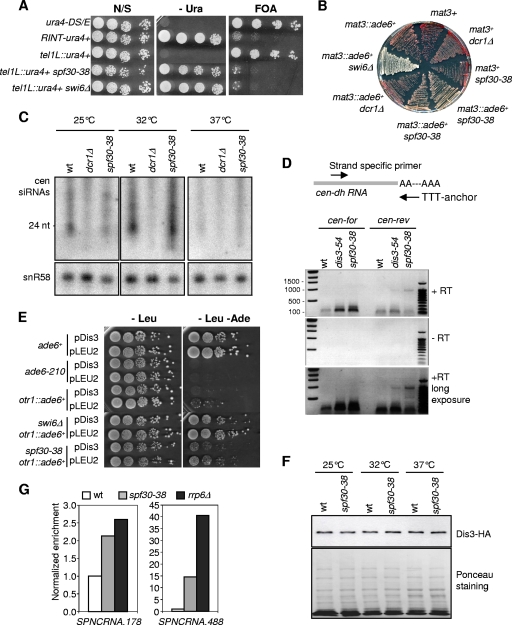
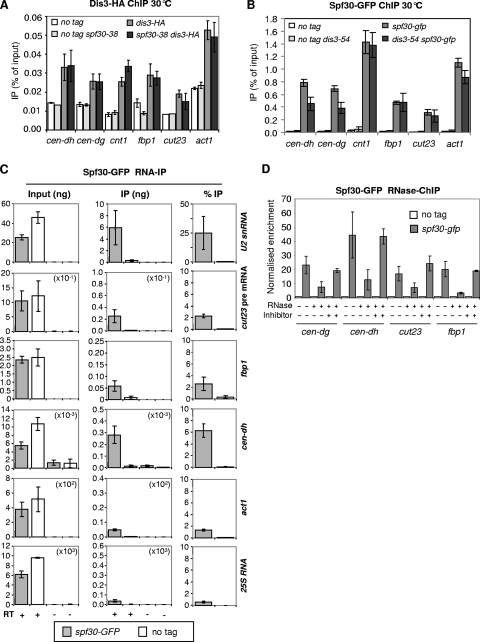
Heterochromatin assembly in fission yeast relies on the processing of cognate noncoding RNAs by both the RNA interference and the exosome degradation pathways. Recent evidence indicates that splicing factors facilitate the cotranscriptional processing of centromeric transcripts into small interfering RNAs (siRNAs). In contrast, how the exosome contributes to heterochromatin assembly and whether it also relies upon splicing factors were unknown. We provide here evidence that fission yeast Spf30 is a splicing factor involved in the exosome pathway of heterochromatin silencing. Spf30 and Dis3, the main exosome RNase, colocalize at centromeric heterochromatin and euchromatic genes. At the centromeres, Dis3 helps recruiting Spf30, whose deficiency phenocopies the dis3-54 mutant: heterochromatin is impaired, as evidenced by reduced silencing and the accumulation of polyadenylated centromeric transcripts, but the production of siRNAs appears to be unaffected. Consistent with a direct role, Spf30 binds centromeric transcripts and locates at the centromeres in an RNA-dependent manner. We propose that Spf30, bound to nascent centromeric transcripts, perhaps with other splicing factors, assists their processing by the exosome. Splicing factor intercession may thus be a common feature of gene silencing pathways.
Figures
Similar articles
-
Splicing factors facilitate RNAi-directed silencing in fission yeast.Science. 2008 Oct 24;322(5901):602-6. doi: 10.1126/science.1164029. Science. 2008. PMID: 18948543 Free PMC article.
-
Involvement of the spliceosomal U4 small nuclear RNA in heterochromatic gene silencing at fission yeast centromeres.J Biol Chem. 2010 Feb 19;285(8):5630-8. doi: 10.1074/jbc.M109.074393. Epub 2009 Dec 14. J Biol Chem. 2010. PMID: 20018856 Free PMC article.
-
Studies on the mechanism of RNAi-dependent heterochromatin assembly.Cold Spring Harb Symp Quant Biol. 2006;71:461-71. doi: 10.1101/sqb.2006.71.044. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381328 Review.
-
Argonaute and Triman generate dicer-independent priRNAs and mature siRNAs to initiate heterochromatin formation.Mol Cell. 2013 Oct 24;52(2):173-83. doi: 10.1016/j.molcel.2013.08.046. Epub 2013 Oct 3. Mol Cell. 2013. PMID: 24095277
-
RITS-connecting transcription, RNA interference, and heterochromatin assembly in fission yeast.Wiley Interdiscip Rev RNA. 2011 Sep-Oct;2(5):632-46. doi: 10.1002/wrna.80. Epub 2011 Mar 23. Wiley Interdiscip Rev RNA. 2011. PMID: 21823226 Free PMC article. Review.
Cited by
-
Splicing of branchpoint-distant exons is promoted by Cactin, Tls1 and the ubiquitin-fold-activated Sde2.Nucleic Acids Res. 2022 Sep 23;50(17):10000-10014. doi: 10.1093/nar/gkac769. Nucleic Acids Res. 2022. PMID: 36095128 Free PMC article.
-
Arabidopsis RRP6L1 and RRP6L2 function in FLOWERING LOCUS C silencing via regulation of antisense RNA synthesis.PLoS Genet. 2014 Sep 11;10(9):e1004612. doi: 10.1371/journal.pgen.1004612. eCollection 2014 Sep. PLoS Genet. 2014. PMID: 25211139 Free PMC article.
-
CPF-associated phosphatase activity opposes condensin-mediated chromosome condensation.PLoS Genet. 2014 Jun 19;10(6):e1004415. doi: 10.1371/journal.pgen.1004415. eCollection 2014 Jun. PLoS Genet. 2014. PMID: 24945319 Free PMC article.
-
Epigenetic Regulation of Chromatin States in Schizosaccharomyces pombe.Cold Spring Harb Perspect Biol. 2015 Jul 1;7(7):a018770. doi: 10.1101/cshperspect.a018770. Cold Spring Harb Perspect Biol. 2015. PMID: 26134317 Free PMC article. Review.
-
A genetic screen for functional partners of condensin in fission yeast.G3 (Bethesda). 2014 Feb 19;4(2):373-81. doi: 10.1534/g3.113.009621. G3 (Bethesda). 2014. PMID: 24362309 Free PMC article.
References
-
- Allshire, R. C., E. R. Nimmo, K. Ekwall, J. P. Javerzat, and G. Cranston. 1995. Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev. 9:218-233. - PubMed
-
- Andrulis, E. D., J. Werner, A. Nazarian, H. Erdjument-Bromage, P. Tempst, and J. T. Lis. 2002. The RNA processing exosome is linked to elongating RNA polymerase II in Drosophila. Nature 420:837-841. - PubMed
-
- Bahler, J., J. Q. Wu, M. S. Longtine, N. G. Shah, A. McKenzie III, A. B. Steever, A. Wach, P. Philippsen, and J. R. Pringle. 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14:943-951. - PubMed
-
- Bentley, D. L. 2005. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr. Opin. Cell Biol. 17:251-256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases