

Phosphoglucomutase of Yersinia pestis is required for autoaggregation and polymyxin B resistance
- PMID: 20028810
- PMCID: PMC2825912
- DOI: 10.1128/IAI.00997-09
Phosphoglucomutase of Yersinia pestis is required for autoaggregation and polymyxin B resistance
Abstract
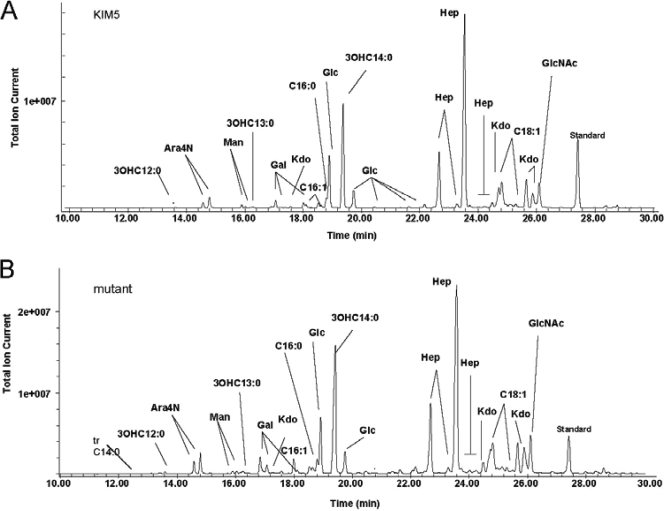
Yersinia pestis, the causative agent of plague, autoaggregates within a few minutes of cessation of shaking when grown at 28 degrees C. To identify the autoaggregation factor of Y. pestis, we performed mariner-based transposon mutagenesis. Autoaggregation-defective mutants from three different pools were identified, each with a transposon insertion at a different position within the gene encoding phosphoglucomutase (pgmA; y1258). Targeted deletion of pgmA in Y. pestis KIM5 also resulted in loss of autoaggregation. Given the previously defined role for phosphoglucomutase in antimicrobial peptide resistance in other organisms, we tested the KIM5 DeltapgmA mutant for antimicrobial peptide sensitivity. The DeltapgmA mutant displayed >1,000-fold increased sensitivity to polymyxin B compared to the parental Y. pestis strain, KIM5. This sensitivity is not due to changes in lipopolysaccharide (LPS) since the LPSs from both Y. pestis KIM5 and the DeltapgmA mutant are identical based on a comparison of their structures by mass spectrometry (MS), tandem MS, and nuclear magnetic resonance analyses. Furthermore, the ability of polymyxin B to neutralize LPS toxicity was identical for LPS purified from both KIM5 and the DeltapgmA mutant. Our results indicate that increased polymyxin B sensitivity of the DeltapgmA mutant is due to changes in surface structures other than LPS. Experiments with mice via the intravenous and intranasal routes did not demonstrate any virulence defect for the DeltapgmA mutant, nor was flea colonization or blockage affected. Our findings suggest that the activity of PgmA results in modification and/or elaboration of a surface component of Y. pestis responsible for autoaggregation and polymyxin B resistance.
Figures
References
-
- Arellano-Reynoso, B., N. Lapaque, S. Salcedo, G. Briones, A. E. Ciocchini, R. Ugalde, E. Moreno, I. Moriyon, and J. P. Gorvel. 2005. Cyclic beta-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat. Immunol. 6:618-625. - PubMed
-
- Arnold, T. M., G. N. Forrest, and K. J. Messmer. 2007. Polymyxin antibiotics for gram-negative infections. Am. J. Health Syst. Pharm. 64:819-826. - PubMed
-
- Bizzini, A., P. Majcherczyk, S. Beggah-Moller, B. Soldo, J. M. Entenza, M. Gaillard, P. Moreillon, and V. Lazarevic. 2007. Effects of alpha-phosphoglucomutase deficiency on cell wall properties and fitness in Streptococcus gordonii. Microbiology 153:490-498. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
