

The small subunit of snapdragon geranyl diphosphate synthase modifies the chain length specificity of tobacco geranylgeranyl diphosphate synthase in planta
- PMID: 20028839
- PMCID: PMC2814502
- DOI: 10.1105/tpc.109.071282
The small subunit of snapdragon geranyl diphosphate synthase modifies the chain length specificity of tobacco geranylgeranyl diphosphate synthase in planta
Abstract
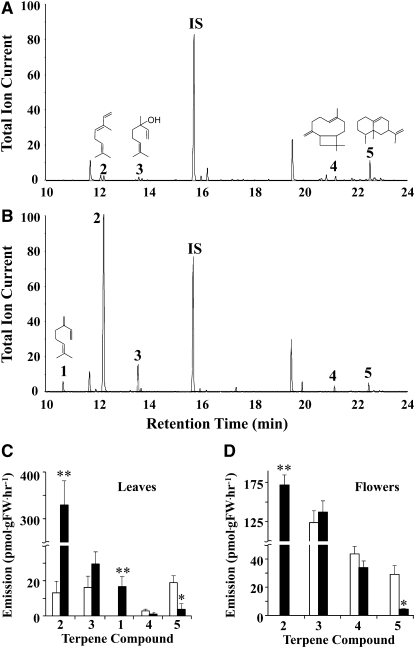
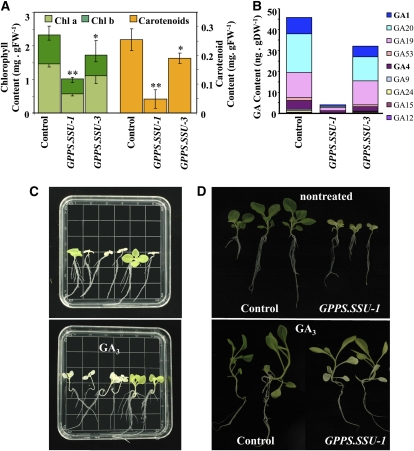
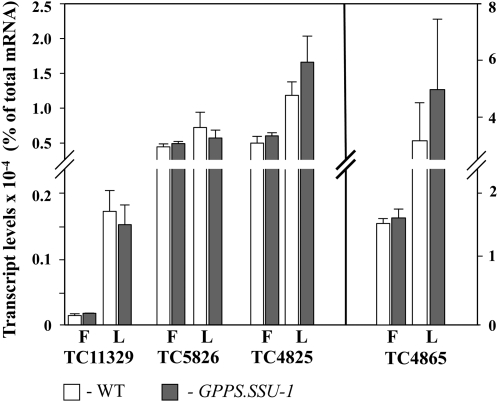
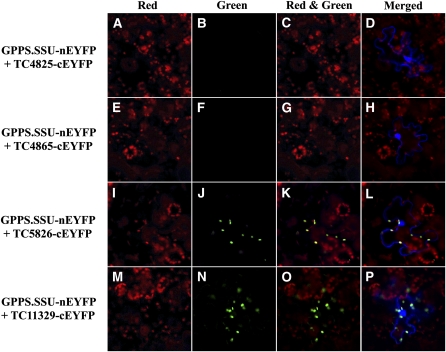
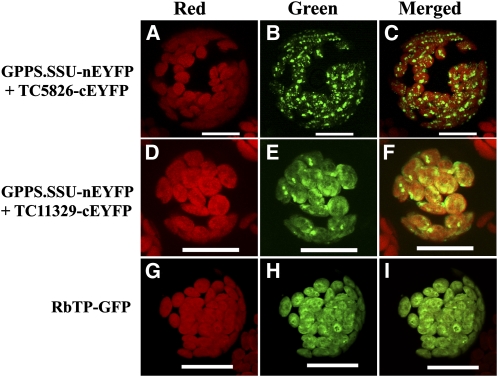
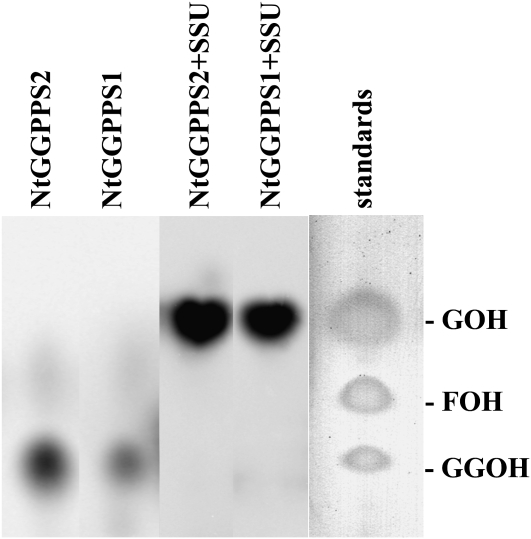
Geranyl diphosphate (GPP), the precursor of many monoterpene end products, is synthesized in plastids by a condensation of dimethylallyl diphosphate and isopentenyl diphosphate (IPP) in a reaction catalyzed by homodimeric or heterodimeric GPP synthase (GPPS). In the heterodimeric enzymes, a noncatalytic small subunit (GPPS.SSU) determines the product specificity of the catalytic large subunit, which may be either an active geranylgeranyl diphosphate synthase (GGPPS) or an inactive GGPPS-like protein. Here, we show that expression of snapdragon (Antirrhinum majus) GPPS.SSU in tobacco (Nicotiana tabacum) plants increased the total GPPS activity and monoterpene emission from leaves and flowers, indicating that the introduced catalytically inactive GPPS.SSU found endogenous large subunit partner(s) and formed an active snapdragon/tobacco GPPS in planta. Bimolecular fluorescence complementation and in vitro enzyme analysis of individual and hybrid proteins revealed that two of four GGPPS-like candidates from tobacco EST databases encode bona fide GGPPS that can interact with snapdragon GPPS.SSU and form a functional GPPS enzyme in plastids. The formation of chimeric GPPS in transgenic plants also resulted in leaf chlorosis, increased light sensitivity, and dwarfism due to decreased levels of chlorophylls, carotenoids, and gibberellins. In addition, these transgenic plants had reduced levels of sesquiterpene emission, suggesting that the export of isoprenoid intermediates from the plastids into the cytosol was decreased. These results provide genetic evidence that GPPS.SSU modifies the chain length specificity of phylogenetically distant GGPPS and can modulate IPP flux distribution between GPP and GGPP synthesis in planta.
Figures

References
-
- Aharoni, A., Jongsma, M.A., and Bouwmeester, H.J. (2005). Volatile science? Metabolic engineering of terpenoids in plants. Trends Plant Sci. 10 594–602. - PubMed
-
- Aharoni, A., Jongsma, M.A., Kim, T., Ri, M., Giri, A.P., Verstappen, F.W., Schwab, W., and Bouwmeester, H.J. (2006). Metabolic engineering of terpenoid biosynthesis in plants. Phytochem. Rev. 5 49–58.
-
- Ament, K., Van Schie, C.C., Bouwmeester, H.J., Haring, M.A., and Schuurink, R.C. (2006). Induction of a leaf specific geranylgeranyl pyrophosphate synthase and emission of (E,E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene in tomato are dependent on both jasmonic acid and salicylic acid signaling pathways. Planta 224 1197–1208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
