

Mycobacterium tuberculosis cAMP receptor protein (Rv3676) differs from the Escherichia coli paradigm in its cAMP binding and DNA binding properties and transcription activation properties
- PMID: 20028978
- PMCID: PMC2844151
- DOI: 10.1074/jbc.M109.047720
Mycobacterium tuberculosis cAMP receptor protein (Rv3676) differs from the Escherichia coli paradigm in its cAMP binding and DNA binding properties and transcription activation properties
Abstract
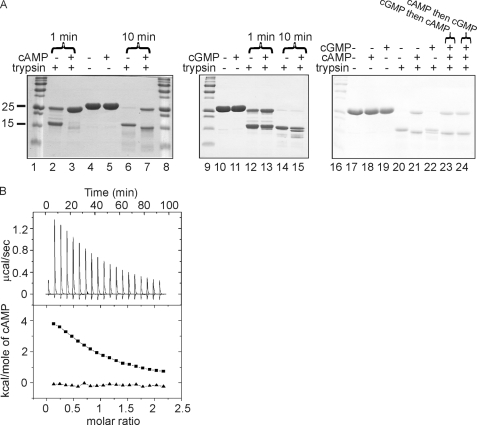
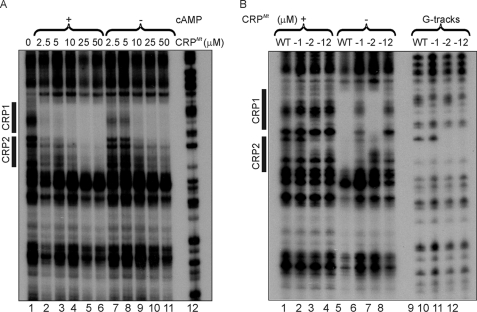
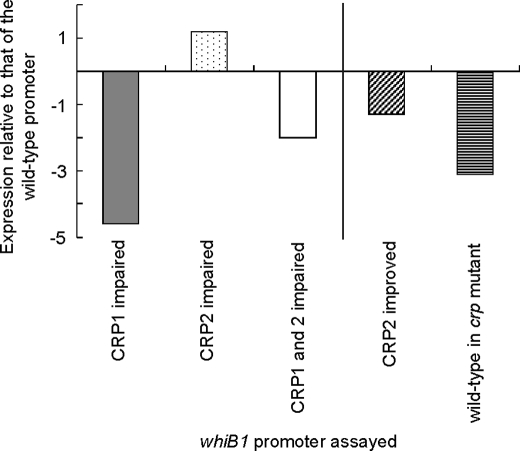
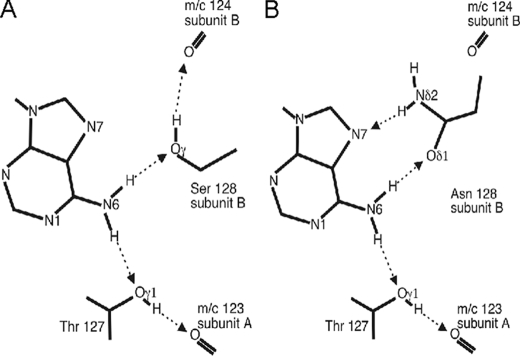
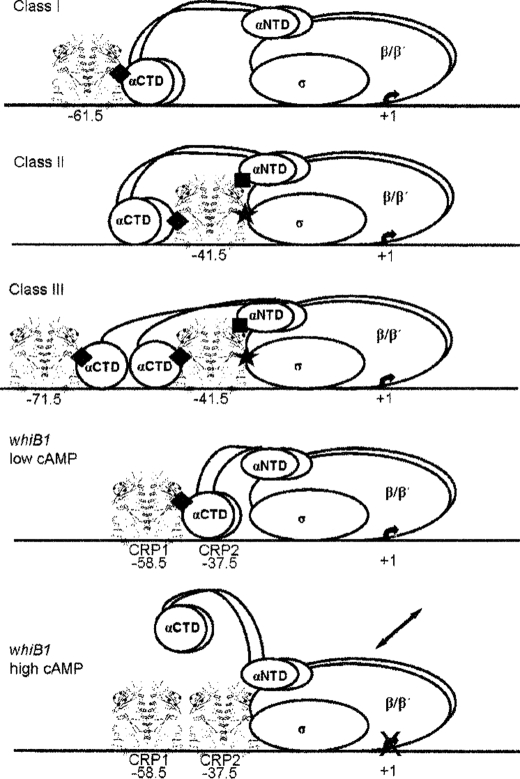
The pathogen Mycobacterium tuberculosis produces a burst of cAMP upon infection of macrophages. Bacterial cyclic AMP receptor proteins (CRP) are transcription factors that respond to cAMP by binding at target promoters when cAMP concentrations increase. Rv3676 (CRP(Mt)) is a CRP family protein that regulates expression of genes (rpfA and whiB1) that are potentially involved in M. tuberculosis persistence and/or emergence from the dormant state. Here, the CRP(Mt) homodimer is shown to bind two molecules of cAMP (one per protomer) at noninteracting sites. Furthermore, cAMP binding by CRP(Mt) was relatively weak, entropy driven, and resulted in a relatively small enhancement in DNA binding. Tandem CRP(Mt)-binding sites (CRP1 at -58.5 and CRP2 at -37.5) were identified at the whiB1 promoter (PwhiB1). In vitro transcription reactions showed that CRP1 is an activating site and that CRP2, which was only occupied in the presence of cAMP or at high CRP(Mt) concentrations in the absence of cAMP, is a repressing site. Binding of CRP(Mt) to CRP1 was not essential for open complex formation but was required for transcription activation. Thus, these data suggest that binding of CRP(Mt) to the PwhiB1 CRP1 site activates transcription at a step after open complex formation. In contrast, high cAMP concentrations allowed occupation of both CRP1 and CRP2 sites, resulting in inhibition of open complex formation. Thus, M. tuberculosis CRP has evolved several distinct characteristics, compared with the Escherichia coli CRP paradigm, to allow it to regulate gene expression against a background of high concentrations of cAMP.
Figures



Similar articles
-
Characterization of Mycobacterium tuberculosis Rv3676 (CRPMt), a cyclic AMP receptor protein-like DNA binding protein.J Bacteriol. 2005 Nov;187(22):7795-804. doi: 10.1128/JB.187.22.7795-7804.2005. J Bacteriol. 2005. PMID: 16267303 Free PMC article.
-
A member of the cAMP receptor protein family of transcription regulators in Mycobacterium tuberculosis is required for virulence in mice and controls transcription of the rpfA gene coding for a resuscitation promoting factor.Mol Microbiol. 2005 Jun;56(5):1274-86. doi: 10.1111/j.1365-2958.2005.04609.x. Mol Microbiol. 2005. PMID: 15882420 Free PMC article.
-
Dysregulation of serine biosynthesis contributes to the growth defect of a Mycobacterium tuberculosis crp mutant.Mol Microbiol. 2011 Oct;82(1):180-98. doi: 10.1111/j.1365-2958.2011.07806.x. Epub 2011 Sep 8. Mol Microbiol. 2011. PMID: 21902733 Free PMC article.
-
Syn, anti, and finally both conformations of cyclic AMP are involved in the CRP-dependent transcription initiation mechanism in E. coli lac operon.Cell Biochem Funct. 2008 Jun;26(4):399-405. doi: 10.1002/cbf.1462. Cell Biochem Funct. 2008. PMID: 18338329 Review.
-
Cyclic-AMP and bacterial cyclic-AMP receptor proteins revisited: adaptation for different ecological niches.Curr Opin Microbiol. 2014 Apr;18(100):1-7. doi: 10.1016/j.mib.2014.01.003. Epub 2014 Feb 7. Curr Opin Microbiol. 2014. PMID: 24509484 Free PMC article. Review.
Cited by
-
Crp is a global regulator of antibiotic production in streptomyces.mBio. 2012 Dec 11;3(6):e00407-12. doi: 10.1128/mBio.00407-12. mBio. 2012. PMID: 23232715 Free PMC article.
-
Cyclic AMP is a critical mediator of intrinsic drug resistance and fatty acid metabolism in M. tuberculosis.Elife. 2023 Feb 22;12:e81177. doi: 10.7554/eLife.81177. Elife. 2023. PMID: 36810158 Free PMC article.
-
Mycobacterium tuberculosis WhiB1 is an essential DNA-binding protein with a nitric oxide-sensitive iron-sulfur cluster.Biochem J. 2010 Dec 15;432(3):417-27. doi: 10.1042/BJ20101440. Biochem J. 2010. PMID: 20929442 Free PMC article.
-
Mycobacterium tuberculosis WhiB1 represses transcription of the essential chaperonin GroEL2.Tuberculosis (Edinb). 2012 Jul;92(4):328-32. doi: 10.1016/j.tube.2012.03.001. Epub 2012 Mar 29. Tuberculosis (Edinb). 2012. PMID: 22464736 Free PMC article.
-
Structure of a Wbl protein and implications for NO sensing by M. tuberculosis.Nat Commun. 2017 Dec 22;8(1):2280. doi: 10.1038/s41467-017-02418-y. Nat Commun. 2017. PMID: 29273788 Free PMC article.
References
-
- Dye C., Scheele S., Dolin P., Pathania V., Raviglione M. C. (1999) J. Am. Med. Assoc. 282, 677–686 - PubMed
-
- Russell D. G. (2001) Nat. Rev. Mol. Cell Biol. 2, 569–577 - PubMed
-
- Stewart G. R., Robertson B. D., Young D. B. (2003) Nat. Rev. Microbiol. 1, 97–105 - PubMed
-
- Cole S. T., Brosch R., Parkhill J., Garnier T., Churcher C., Harris D., Gordon S. V., Eiglmeier K., Gas S., Barry C. E., 3rd, Tekaia F., Badcock K., Basham D., Brown D., Chillingworth T., Connor R., Davies R., Devlin K., Feltwell T., Gentles S., Hamlin N., Holroyd S., Hornsby T., Jagels K., Krogh A., McLean J., Moule S., Murphy L., Oliver K., Osborne J., Quail M. A., Rajandream M. A., Rogers J., Rutter S., Seeger K., Skelton J., Squares R., Squares S., Sulston J. E., Taylor K., Whitehead S., Barrell B. G. (1998) Nature 393, 537–544 - PubMed
-
- Agarwal N., Lamichhane G., Gupta R., Nolan S., Bishai W. R. (2009) Nature 460, 98–102 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
